
ARTIGO DE PESQUISA
Respostas fisiológicas de Theobroma cacao L.submetidas ao alagamento por longos períodos
Physiological responses of Theobroma cacao L. subjected to flooding for long periods
https://doi.org/10.32712/2446-4775.2024.1325
- Silva, Bruno Eustáquio Cirilo1*
 https://orcid.org/0000-0002-2840-2859
https://orcid.org/0000-0002-2840-2859
- Silva, Marlinda Rufina Jolomba2
https://orcid.org/0000-0002-9098-2479
- 1Universidade Federal do Espírito Santo (UFES), Campus São Mateus – CEUNES, BR-101, km 60, Litorâneo, CEP 29932-540, São Mateus, ES, Brasil.
- 2Universidade Federal do Sul da Bahia (UFBA), Centro de Formação em Ciências e Tecnologias Agroflorestais. Ceplac, Centro, CEP 45653-919, Ilhéus, BA, Brasil.
- *Correspondência:
- brunomukuri@gmail.com
Resumo
O trabalho teve como objetivo avaliar respostas fisiológicas de plantas de cacau (Theobroma cacao L.) submetidas ao estresse hídrico sob alagamento em diferentes períodos. O trabalho foi conduzido sob casa de vegetação no Instituto Capixaba de Pesquisa, Assistência Técnica e Extensão Rural (INCAPER), localizado em linhares-Es. cultivares utilizadas de Theobroma cacao L. foram variedades de cacau CCN 51 e TSH 1188, sendo a primeira utilizada como copa e a segunda como porta enxerto. O experimento foi instalado com mudas de cacau sob estresse em ambiente alagado, dispostas em baldes e submersas de água, sob condições de tempos diferentes. As leituras foram realizadas aos 0, 1, 2, 4 e 8 dias de alagamento. Foram realizadas medidas assimilação liquida de CO2, condutância estomática, transpiração e eficiência do uso de água, todos utilizando o IRGA. O tratamento de 2 dias após o alagamento não diferenciou do tratamento controle (sem alagamento), enquanto no tratamento de 4 DAA houve uma redução no teor de assimilação liquida de CO2. As plantas alagadas por 8 dias apresentaram crescimento de lenticelas, mostrando a tentativa de amenizar as condições ambientais. Não houve interferência pelo alagamento nos fotossistemas PI e PII o aparato fotossintético se manteve estável.
- Palavras-chave:
- Alagamento.
- Rendimento fosossintético.
- Fluorescência.
Abstract
The aim of this study was to evaluate the physiological responses of cocoa plants (Theobroma cacao L.) subjected to water stress under flooding in different periods. The study was conducted in a greenhouse at the Capixaba Institute for Research, Technical Assistance and Rural Extension (INCAPER), located in Linhares-Es. The cultivars used for Theobroma cacao L. were cocoa varieties CCN 51 and TSH 1188, the first being used as a scion and the second as a rootstock. The experiment was carried out with cocoa seedlings under stress in a flooded environment, arranged in buckets and submerged in water, under different time conditions. Readings were taken at 0, 1, 2, 4 and 8 days of flooding. Measurements of net CO2 assimilation, stomatal conductance, transpiration and water use efficiency were performed, all using IRGA. The treatment 2 days after flooding did not differ from the control treatment (without flooding), while in the treatment 4 DAA there was a reduction in the net CO2 assimilation content. Plants flooded for 8 days showed lenticel growth, showing an attempt to alleviate environmental conditions. There was no interference by flooding in the PI and PII photosystems; the photosynthetic apparatus remained stable.
- Keywords:
- Flooding.
- Phosynthetic yield.
- Fluorescence.
Introdução
O cacau (Theobroma cacao L.) é originário do Norte do Brasil (Amazônia). A produção de lavouras de cacau atingiu o ápice na década de 1980, onde o país se tornou um dos maiores produtores mundiais do fruto, com destaque para a produção no estado da Bahia[1]. No estado do Espírito Santo a produção de cacau é concentrada na área de Linhares, onde com o objetivo de alcançar competitividade e reconhecimento no mercado tem-se buscado melhorar aspectos de produção[2]. O município de maior produção no estado do Espírito Santo é o de Linhares com cerca de 88% da área planta do estado[3]. Na região norte do Espírito Santo, onde está localizado o município de maior produção, a maior parte da região cacaueira é composta de solos aluviais e sujeita ao alagamento, principalmente no verão[4].
Inundações frequentes podem reduzir a produtividade de muitas culturas, o que torna essencial a identificação de cultivares tolerantes à inundação, de modo a garantir a produtividade mesmo quando as regiões produtoras de cacau são atingidas pela água causando o alagamento. Ainda de acordo com Braga[4], plantas tolerantes ao estresse por alagamento dependem da espécie, da duração do estresse a qual foi submetida, o estágio de desenvolvimento em que a planta se encontra e das condições ambientais. O efeito do alagamento no sistema radicular, em plantas terrestres, pode ser prejudicial ou mesmo letal, pois bloqueia a transferência de oxigênio e outros gases entre o solo e a atmosfera[5,6]. As respostas das plantas em condições de alagamento podem incluir o fechamento estomático, senescência foliar prematura, redução do volume de raízes e do crescimento da parte aérea[7,8].
Diminuição em relação ao crescimento, área foliar, condutância estomática, taxas fotossintéticas, somados com a formação das lenticelas e raízes adventícias colaboram para que a planta consiga lidar com as condições anaeróbias[7,8,4]. Em condições de alagamento as plantas reduzem as trocas gasosas; a absorção de nutrientes, o volume das raízes, altera o armazenamento e translocação de fotossintetizados, e induz a produção de hormônios endógenos, tais como etileno[9,4]. Nessas condições os microrganismos do solo são forçados a reduzir receptores de elétrons diferentes de O2, consequentemente o potencial redox do solo cai[4].
O crescimento das plantas de cacau é inibido quando submetido a solos alagados reduzindo assim a produção dos frutos, devido ao aumento do tempo do período da primeira floração e reduzir o crescimento vegetativo[10,4]. Ao avaliarem seis clones de cacau Rehem et al.[11] verificaram que as taxas fotossintéticas foram diferentes em comparação ao ambiente não alagado.
O objetivo do presente estudo foi analisar as respostas das plantas de cacau submetidas ao estresse por alagamento em diferentes períodos.
Materiais e Métodos
O experimento foi conduzido em casa de vegetação, sob condições naturais. Conduzido no Instituto Capixaba de Pesquisa, Assistência Técnica e Extensão Rural (INCAPER), localizado no município de Linhares no norte do Estado do Espírito Santo. As cultivares utilizadas de Theobroma cacao L foram variedades de cacau CCN 51 e TSH 1188, sendo a primeira utilizada como copa e a segunda como porta enxerto. O experimento foi instalado com mudas de cacau sob estresse em ambiente alagado, dispostas em baldes e submersas de água, sob condições de tempos diferentes. As leituras foram realizadas aos 0, 1, 2, 4 e 8 dias de alagamento (FIGURA 1).
FIGURA 1: Plantas submetidas ao estresse por alagamento.
Foram realizadas medidas assimilação liquida de CO2 (A), condutância estomática (gs), transpiração (E) e eficiência do uso de água, todos utilizando o IRGA (Infra-red Gas Analyzer) (FIGURA 2). Essas variáveis foram medidas durante o período entre 09h e 12h em todas as plantas. Nas medidas foram utilizadas folhas completamente expandidas, expostas ao sol durante todo período de medidas, segundo Machado et al.[12] e em condições de estresse por alagamento.
FIGURA 2: Aparelho IRGA (A) e utilização do aparelho (B).
Foram efetuadas concomitantemente com as da fotossíntese nas plantas avaliadas, com a utilização de um fluorômetro portátil (Handy-PEA, Hansatech Instruments Ltd., King's Lynn Norfolk, UK) (FIGURA 3), induzida pela luz vermelha (pico em 650 nm) de aproximadamente 3.000 µmol m-2 s-1, fornecida por um conjunto de três diodos emissores de luz, e gravados por 1s com resolução de 12 bit. 17 2.4 Teste-JIP A fluorescência transiente da clorofila foi analisada de acordo com o Teste-JIP (STRASSER; SRIVASTAVA; TSIMILLI-MICHAEL, 500; STRASSER; TSIMILLIMICHAEL; SRIVASTAVA, 2004; TSIMILLI-MICHAEL; STRASSER, 2008), utilizando as intensidades de fluorescência medidas a: 50 µs (fluorescência inicial – F0), 2 ms (fluorescência no ponto J – FJ), 30 ms (fluorescência no ponto I – FI), e a fluorescência máxima (FM). Utilizaram-se três plantas de cada tratamento, associação do porta-enxertos TSH 1188 com a copa CNN 51, totalizando 15 plantas, sendo feitas duas medidas por planta. As medidas de fluorescência da clorofila foram realizadas em folhas intactas, ligadas à planta, e adaptadas previamente ao escuro por 30 minutos. A partir destas intensidades, foram calculados os fluxos específicos por centro de reação, eficiência ou rendimentos quânticos, e índices de desempenho.
FIGURA 3: Medições utilizando o aparelho Handy-PEA.
O delineamento experimental utilizado fui o inteiramente cazualizado(DIC). Os dados amostrais foram submetidos ao Software Estatístico Livre R e ao Microsoft Excel, sendo utilizado o Teste Tukey a 5% de probabilidade.
Resultados e Discussão
O crescimento de lenticelas hipertróficas nas plantas de cacau (FIGURA 4) foram observadas, após 8 dias de alagamento. Kozlowski[7] observou que resposta a adaptação primária das plantas ao alagamento do substrato é a capacidade de absorver O2 pelos tecidos aéreos, aumentando sua concentração nesses tecidos e favorecendo as formações de lenticelas hipertróficas, aerênquima e raízes adventícias. A clorose não foi observada nas folhas de nenhuma planta. De acordo com Bertolde et al.[13] a clorose só foi observada nas plantas submetidas ao alagamento após 15 dias.
FIGURA 4: Lenticelas nas plantas de cacau após 8 dias de alagamento.
Nas plantas com 4 e 8 dias (FIGURA 5) epinastia foliar induzidas pelo alagamento, em condições de alagamento ou anaeróbicas ao redor das raízes induz o aumento da síntese de etileno na parte aérea da planta, conduzindo a resposta epinástica.
FIGURA 5: Plantas com epinastia foliar induzidas pelo alagamento.
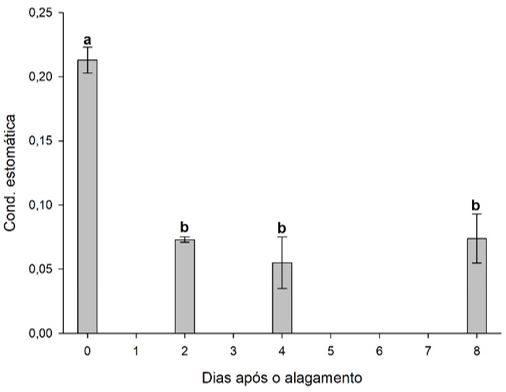
Conforme observado abaixo, na TABELA 1, a assimilação liquida de CO2 (A) das mudas de cacaueiro houve influência pelo alagamento das plantas (FIGURA 6). O tratamento de 2 dias após o alagamento não diferenciou do tratamento controle (sem alagamento), enquanto no tratamento de 4 DAA houve uma redução no teor de assimilação liquida de CO2, inicialmente, a diminuição de A é devida ao fechamento estomático, resultando na diminuição da absorção de CO2 pelas folhas[14,7]. O fechamento de estômatos sob condições inundadas pode estar relacionado a uma diminuição na condutividade hidráulica da raiz[15,16], promovendo uma diminuição em gs (FIGURA 7).
| Tratamentos | A | gs | E | EUA |
|---|---|---|---|---|
| 0 DAA | 10.27 a | 0.210 a | 3.64 a | 2.84 a |
| 2 DAA | 5.39 ab | 0.0729 b | 1.55 b | 3.50 a |
| 4 DAA | 3.57 b | 0.0556 b | 1.45 b | 3.03 a |
| 8 DAA | 5.21 ab | 0.0738 b | 1.12 b | 3.58 a |
| CV (%) | 35.13 | 24.86 | 22.28 | 22.92 |
| Médias seguidas de mesma letra na coluna não diferem entre si pelo teste de Tuckey (<0,05). | ||||
Em árvores frutíferas, o declínio nas taxas de transpiração (E) em resposta ao alagamento (FIGURA 8) é provavelmente devido a uma diminuição de gs, uma vez que a deficiência de O2 não diminuiu significativamente o potencial hídrico do xilema[17]. Não foi observada diferença significativa para EUA, corroborando com trabalhos de Mielke et al.[18], onde não se observou significância de EUA em Genipa americana e, também, com trabalho de Bertolde et al.[13] com Theobroma cacao L.
O estresse por alagamento promove a redução das taxas transpiratórias das plantas, resultante das mudanças na condutância estomática, pois, nessa situação, a rota apoplástica poderá ser pouco usada e a resistência hidráulica, consequentemente, aumentada[19,20]. Dessa forma, a avaliação periódica de parâmetros fisiológicos, durante a exposição de uma determinada espécie vegetal ao estresse ambiental, permite uma melhor compreensão dos mecanismos específicos de respostas aos fatores estressantes e de sua evolução no tempo[21].


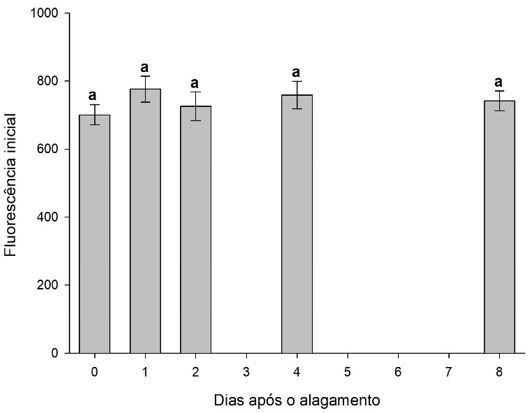
| Tratamentos | Fo | Fm | Fv/Fm | Fv/Fo | PI(ABS) | SPAD |
|---|---|---|---|---|---|---|
| 0 DAA | 700.67 a | 3778.17 a | 0.77 a | 3.44 a | 13.14 a | 38.00 a |
| 1 DAA | 766.33 a | 3904.17 a | 0.74 a | 2.90 a | 6.86 a | 36.48 a |
| 2 DAA | 725.83 a | 3477.67 a | 0.74 a | 3.01 a | 10.95 a | 41.63 a |
| 4 DAA | 758.50 a | 3348.50 a | 0.67 a | 2.45 a | 5.58 a | 38.97 a |
| 8 DAA | 741.67 a | 3486.00 a | 0.72 a | 2.68 a | 5.78 a | 37.57 a |
| CV (%) | 12.04 | 16.42 | 11.01 | 22.97 | 58.51 | 15.13 |
| Médias seguidas de mesma letra na coluna não diferem entre si pelo teste de Tuckey (<0,05). | ||||||
Os valores de Fv/Fm (FIGURA 9, 10 e 11) são quase constantes (cerca de 0,832 ± 0,004) para a maioria das espécies de plantas sob condições não limitativas[22]; no entanto, em plantas estressadas, valores menores indicam a ocorrência de fotoinibição ou outras lesões no complexo de coleta de luz do PSII[23]. Além disso, plantas estressadas mostraram um aumento de Fo, indicando uma diminuição do fluxo de elétrons através do PSII[24]. Bertolde et al.[25], trabalhando com 35 clones de T. cacao sob condições inundadas, mostraram decréscimo na relação Fv/Fm e aumento nos valores de Fo para algumas variedades clonais, como observado nos trabalhos de Parolin[26], Waldhoff et al.[27], e Rengifo et al.[28] os valores de FV/FM não demonstraram sensibilidade para detectar os efeitos do alagamento, exceto sob condições extremas[27].


Conforme observado na (TABELA 2), o alagamento não influenciou significativamente (FIGURA 12). Resultado semelhante foi encontrado por Martinazo et al.[29], onde índice de performance (PIABS), atingindo numericamente os menores valores ao final do ensaio. Entretanto, para as plantas submetidas ao alagamento, observou-se diminuição no segundo dia de estresse, seguido com o retorno para os valores do controle no quarto dia, com subsequente redução até o décimo dia.

Conforme verificado na TABELA 2, o alagamento não influenciou no índice SPAD (FIGURA 13), devido ao curto período do alagamento, resultados semelhantes também encontrados por (Mercado Luna et al.[30].

Conclusão
As plantas alagadas por 8 dias apresentaram o crescimento de lenticelas, mostrando a tentativa de amenizar as condições ambientais; não houve interferência pelo alagamento nos fotossistemas PI e PII o aparato fotossintético se manteve estável; O índice SPAD se manteve igual para todas as plantas avaliadas; as plantas sem alagamento mostraram melhores índices de rendimento fotossintéticos.
Fontes de Financiamento
Não houve apoio financeiro.
Conflito de Interesses
Não há conflito de interesses.
Agradecimentos
Ao Instituto Capixaba de Pesquisa Assistência Técnica e Extensão Rural (INCAPER) e à Universidade Federal do Espírito Santo, pelo uso da estrutura e equipamentos.
Colaboradores
Concepção do estudo: BECS; MRJS
Curadoria dos dados: BECS, MRJS
Coleta de dados: BECS
Análise dos dados: BECS
Redação do manuscrito original: BECS; MRJS
Redação da revisão e edição: BECS; MRJS.
Referências
1. Hansen DS. Cultura do cacau. 2017.
2. Teixeira Neto E, Pigatto GA, Lourenzani AEBS. Produção de cacau com Indicação Geográfica (IG) na região de Linhares/ES: aspectos de governança e qualidade. Rev Bras Gestão Desenv Reg. 2018; 14(3): 194-2018. [https://www.rbgdr.net/revista/index.php/rbgdr/article/view/3811/691].
3. Brasil. Ministério da Economia. Instituto Brasileiro de Geografia e Estatística - IBGE. Banco de dados agregados. SIDRA. [acesso em: 29 jun. 2018]. Disponível em: [https://ww2.ibge.gov.br/home/estatistica/indicadores/agropecuaria/lspa/defaulttab.shtm].
4. Braga PCS. Respostas fisiológicas de Theobroma cacao L. a diferentes concentrações de ferro sob alagamento. Vitória. 2016. Dissertação de Mestrado. [Programa de Pós-Graduação em Biologia Vegetal]. Universidade Federal do Espírito Santo, UFES, Vitória. ES. 2016. [https://bdtd.ibict.br/vufind/Record/UFES_92c451ce8b8e983182e81c4eef8878f7].
5. Drew MC. Oxygen deficiency and root metabolism: injury and acclimation under hypoxia and anoxia. Annual Rev Pl Biol. 1997; 48(1): 223-250. [https://doi.org/10.1146/annurev.arplant.48.1.223].
6. Perbonii EGMAT, De Oliveira PV, Bacarini VJBM. A. Atividade fotossintética em plantas de ameixeira submetidas ao déficit hídrico e ao alagamento. Ciênc Rural. 2013; 43(1): [https://doi.org/10.1590/S0103-84782012005000126].
7. Kozlowski TT. Responses of woody plants to flooding and salinity. Tree Physiology Monograph. 1997; 1: 1–29. [https://doi.org/10.1093/treephys/17.7.490].
8. Rehem BC. Respostas fisiológicas de clones de Theobroma cacao L. ao alagamento do substrato. Ilhéus. 2006. 79f. Dissertação de mestrado, Universidade Estadual de Santa Cruz, Ilhéus-BA, 2006.
9. Pezeshki SR. Wetland plant responses to soil flooding. Envir Exper Bot. 2001; 46: 299-312. [https://doi.org/10.1016/S0098-8472(01)00107-1].
10. Sena ARG, Kozlowski TT. The effects of flooding on water relations and growth of Theobroma cacao var. Catongo seedlings. J Horticult Sci. 1986; 61: 265-276. [https://doi.org/10.1080/14620316.1986.11515700].
11. Rehem BC, Almeida AAF, Mielke MS, Gomes FP, Valle RR. Photosynthetic and growth responses of Theobroma cacao L. clones to Water logging. J Trop Agric. 2010; 48: 17-22. [https://www.researchgate.net/publication/275029978_Photosynthetic_and_growth_responses_of_Theobroma_cacao_L_Clones_to_waterlogging].
12. Medri ME, José AEB, Colli S, Muller C. Estudos sobre tolerância ao alagamento em espécies arbóreas nativas da bacia do rio Tibagi. Medri ME, Bianchini E, Shibatta AO, Pimenta JA. A bacia do rio Tibagi. Londrina: Copyright, p. 133-172, 2002.
13. Bertolde FZ, Almeida AAF, Pirovani CP, Gomes FP, Ahnetr D, Baligar VC, et al. Physiological and biochemical responses of Theobroma cacao L. genotypes to flooding. Photosynthetica. 2012; 50(3): 447–457. [https://doi.org/10.1007/s11099-012-0052-4].
14. Pezeshki SR. Differences in patterns of photosynthetic responses to hypoxia in flood-tolerant and flood-sensitive tree species. Photosynthetica. 1993; 28: 423-430.
15. Anderson ME, Gargar ML, Ramsey JC. Inhalation pharmacokinetics: Evaluating systemic extraction, total in vivo metabolism, and the time course of enzyme induction for inhaled styrene in rats based on arterial blood: inhaled air concentration ratios. Toxicol Applied Pharmacol. 1984; 73(1): 176-187. [https://doi.org/10.1016/0041-008X(84)90065-6].
16. Davies FS, Flore JA. Flooding, gas exchange and hydraulic root conductivity of highbush blueberry. Physiol Plantarum. 1986; 67(4): 545-551. [https://doi.org/10.1111/j.1399-3054.1986.tb05053.x].
17. Schaffer B, Anderson PC, Ploetz RC. Responses of fruit crops to flooding. Hort Rev. 1992, 13: 257-313. [https://doi.org/10.1002/9780470650509.ch7].
18. Mielke MS, Almeida AAF, Gomes FP, Aguilar MAG, Mangabeira PAO. Leaf gas exchange, chlorophyll fluorescence and growth responses of Genipa americana seedlings to soil flooding. Envir Exper Bot. 2003; 50: 221–231. [https://doi.org/10.1016/S0098-8472(03)00036-4].
19. Steudle E, Peterson CA. How does water get through roots. J Exper Bot. 1998; 49(322): 775-788. [https://doi.org/10.1093/jxb/49.322.775].
20. Jones HG. Stomatal control of photosynthesis and transpiration. J Exper Bot. 1998; 49: 387-398. [https://doi.org/10.1093/jxb/49.Special_Issue.387].
21. Lichtenthaler HK. Vegetation stress: an introduction to stress concepts in plants. J Pl Physiol. 1996; 148: 4-14. [https://doi.org/10.1016/S0176-1617(96)80287-2
22. Bjorkman O, Demming B. Photon yield de O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta. 1987; 170: 489-504. [https://doi.org/10.1007/BF00402983].
23. Krause GH, Weis E. Chlorophyll fluorescence and photosynthesis: the basis. Annual Review of Plant Physiology and Plant Molecular Biology. 1991; 42: 313-349. [https://doi.org/10.1146/annurev.pp.42.060191.001525].
24. Oliveira HO, Castro GLS, Correa LO, Silvestre WVD, Nascimento SV, Valadares RBS, et al. Coupling physiological analysis with proteomic profile to understand the photosynthetic responses of Young Euterpe oleracea palms to drought. Photosynthesis Res. 2019; 140(2): 189-205. [https://doi.org/10.1007/s11120-018-0597-6].
25. Bertolde FZ, Almeida AAF, Corrêa RX, Gomes FP, Gaiotto FA, Baligar VC et al. Molecular, physiological and morphological analysis of waterlogging tolerance in clonal genotypes of Theobroma cacao L. Tree Physiol. 2010; 30: 56–67. [https://doi.org/10.1093/treephys/tpp101].
26. Parolin P, Junk WJ, Piedade MTF. Gas exchange of six tree species from Central Amazonian floodplains. Tropical Ecol. 2001; 42(1): 15-24. [https://repositorio.inpa.gov.br/handle/1/20928].
27. Waldhoff D, Furch B, Junk WJ. Fluorescence parameters, Chlorophyll concentration, and anatomical features as indicators for flood adaptation of an abundant tree species in Central Amazonia: Symmeria paniculata. Environ Exprimental Botany. 2002; 48: 225-235. [https://doi.org/10.1016/S0098-8472(02)00037-0].
28. Rengifo E, Tezara W, Herrera A. Water relations, Chlorophyll a fluorescence, and contents of saccharides in tree species of a tropical forest in response to flood. Photosynthetica. 2005; 43: 203-210. [https://doi.org/10.1007/s11099-005-0034-x].
29. Martinazzo EG, Perboni AT. Oliveira PV. Bianchi VJ. Bacarin MA. Atividade fotossintética em plantas de ameixeira submetidas ao déficit hídrico e ao alagamento. Ciênc Rural. 2013, 43(1): 35-41. [https://doi.org/10.1590/S0103-84782012005000126].
30. Mercado-Luna A. Nitrogen determination on tomato (Lycopersicon esculentum Mill.) seedlings by color image analysis (RGB). Afric J Biotechnol. Lagos. Aug. 2013; 9(33): 5326-5332. [https://www.ajol.info/index.php/ajb/article/view/92074].