Abstract
Despite human immunodeficiency virus (HIV) latency still being a key obstacle to permanent cure of infection, several synthetic and natural compounds have already been able to efficiently activate HIV-LTR (long terminal repeat), responsible for re-activating the latent virus. In this study, we report that dichloromethane extract of Jatropha gossypiifolia species (JGDM) has dual effect for inhibition of HIV-1 infection and agonist of protein kinase C (PKC). Using MT-4 cells infected with HIV-1 NL4.3, we found that JGDM is able to decrease HIV-1 replication, with an EC50 of 17.96 μg/mL and promotes reactivation of latent provirus in J-Lat 8.4 and 10.6 cells by 4% and 32%, respectively. In addition, the CD4 expression test showed a 66% reduction of CD4 molecules from surface cells. It was also observed that PKC inhibitor, GÖ6983 inhibited the reactivation and CD4 downregulation activity, suggesting that bioactive compound present in the extract acts through the PKC pathway. In conclusion, the dichloromethane extract of J. gossypiifolia showed dual effect with anti-HIV potential, probably by promoting downregulation of CD4 molecules from cell surface, and potential for reactivating latent HIV-1 through PKC pathway.
Natural Products; HIV; Antiviral; Bellyache bush; PKC agonist
Introduction
Over the last two decades, great progress has been made in the fight against human immunodeficiency virus (HIV), such as the use of highly active antiretroviral therapy (HAART), which drastically reduced the viral load to undetectable levels and increased CD4+ T lymphocyte counts in patients living with HIV, resulting in reduced mortality, prevention of progression to acquired immunodeficiency syndrome (AIDS), and increased survival rates of individuals[1,2].
Despite the prolonged suppression of plasma virus by antiretroviral therapy (ART), the major barrier to curing HIV infection are the ‘provirus’ latent reservoirs, responsible for being a source of HIV when treatment is interrupted. ART acts through interference with different stages of HIV replication cycle and only inhibits viruses with active replication activity. Thus, it has little influence on the latent viral reservoirs, making the latency of HIV a fundamental obstacle to a permanent cure of the infection[3].
Several therapeutic approaches are being considered to control or eliminate these latent virus reservoirs[4]. One of the strategies for reservoir elimination is called “shock and kill”, which aims to treat individuals on ART with HIV-activating agents to force viral replication in cells with latent virus reservoirs[5]. Subsequently, the reactivation of HIV expression would induce viral cytopathic effects, immune clearance, and cell death, thus eliminating latently infected cells, while uninfected cells are protected by antiretroviral therapy, blocking de novo infection[6].
Among the approaches studied for the 'shock' of this strategy, the protein kinase C (PKC) pathway has been shown to play an important role in the reactivation of cellular latency through nuclear factor kappa B (NF-κB) signaling, leading to the nuclear translocation of NF-κB and, subsequently, the activation of the HIV-LTR (long terminal repeat)[7]. Several synthetic and natural compounds have already been able to activate PKC isoforms by binding to their regulatory domain, mimicking the diacylglycerol physiological ligand (DAG), such as PEP005 (ingenol-3-angelate)[8], Ingenol-B, isolated from the Euphorbia tirucalli latex (ingenol-3-hexanoate)[9], lactone bryostatin-1, isolated from the marine bryozoan Bugula neritin[10]. However, despite the wide availability of latency reverse agents (LRAs), some compounds, although active, still present some barriers such as toxicity and low potency[11].
Belonging to the Euphorbiaceae family, the genus Jatropha has more than 70 shrub species between which lies the species Jatropha gossypiifolia popularly known as "bellyache bush"[12]. Pharmacological studies have demonstrated significant action of the extract and/or isolated compounds from this plant as antimicrobials, anti-inflammatories, antidiarrheals, antihypertensive and anticancer agents, among others, supporting some of its popular uses[12,13].
Therefore, in the present work, the dichloromethane extract obtained from J. gossypiifolia leaves, collected in the Brazilian Amazon, was tested to verify its in vitro antiviral activity and latent HIV reactivation potential.
Materials and Methods
Plant material and extraction
The leaves of J. gossypiifolia Linneus were collected from Rio Branco, Acre, Brazil (9°58'55.9"S 67°49'52.4"W) in July of 2016. The identification and authentication of the plant was made at the Herbarium of the Zoobotanical Park, Federal University of Acre (UFAC) and a voucher specimen was deposited in the herbarium under registration number UFACPZ20439. The plant material was incubated at 40°C until completely dehydrated and then triturated to 100 g and used for extraction by exhaustive maceration with 500 mL of ethanol (99%) at room temperature. The obtained macerate was vacuum filtered and subjected to the rotary evaporator at 55°C as previously described in literature. The crude alcoholic extract obtained was solubilized on 1:1 methanol and fractionated by liquid-liquid partition with solvents of increasing polarity in order to obtain the hexane (C6H14), dichloromethane (CH2Cl2), ethyl acetate (AcOEt), n-butanol (BuOH) extracts followed by solvent evaporation in rotary evaporator at 55ºC. The hexane, ethyl acetate, and n-butanol extracts mentioned above were tested and found to be non-HIV-reactivating in J-Lat 8.4 cells (data not shown), whereas the dichloromethane extract was found to be HIV-reactivating, warranting further investigation.
Cells and virus
J-Lat (8.4 and 10.6 clones) and MT-4 cells were kindly provided by AIDS Reagent Program (NIAID). J-Lat and MT-4 cells were cultured in RPMI 1640 medium (Life Technologies) supplemented with 10% fetal bovine serum (FBS) at 37°C and 5% CO2 atmosphere. The HIV-1 (NL4.3) virus stock used was obtained by Abreu et al.[9] in a previous work where 293LTV cells were transfected with pNL4.3 (AIDS Reagent Program) using lipofectamine (Life Technologies).
Reagents
PKC inhibitors (GÖ6983, GÖ6976 and RO-31-8220 - Sigma- Aldrich), Ingenol-B (Ing-B - Kyolab) and lamivudine (3TC - AIDS Reagent Program) were used. Ing-B and 3TC were resuspended in dimethyl sulfoxide (DMSO) to 200 mM concentration and stored in -80°C freezer. PKC inhibitors were resuspended in DMSO at 10 mM concentration and stored in -20ºC freezer.
Cytotoxic effect assay
MT-4 cells were seeded in 96-well microplates at a density of 104 cells per well and incubated with 2-fold serial diluted concentrations of J. gossypiifolia dichrolomethane (JGDM) extract (2.5 to 80 μg/mL) or 3TC (0.375 to 48 μg/mL) at 37°C and atmosphere 5% CO2 for 5 days. DMSO-treated cells (mock) were also incubated and used as positive control. Cell viability was determined using the CellTiter-blue fluorescence Cell Viability Assay (Promega) following the manufacturer’s instructions. Briefly, 20 µL of CellTiter was added to each well followed by incubation overnight. After 24 hours, fluorescence was determined on a Spectramax M2 (Molecular Devices) at 560ex/590em nm.
Anti-HIV-1 activity assay
MT-4 cells were infected with HIV-1 NL4.3 clone (MOI = 0.002) by spinoculation for 2 hours at 1200 x g and then seeded in 96-well microplates at a density of 104 cells per well. Uninfected cells (mock) were also submitted to the spinoculation process. After seeding, JGDM extract was added following a 2-fold serial diluation (2.5 to 80 μg/mL) and incubation at 37°C and 5% CO2. Because HIV-1 NL4.3 infection leads to cell death, antiviral activity was distinguished from potential cell death caused by HIV-1 infectivity based on cell viability. HIV infected cells without extract treatment and mock were used to normalize cell viability values. 3TC (0.375 to 48 μg/mL), previously used by our group, was used as positive control[14]. Five days after infection and treatment with JDM extract, cell viability was measured by adding CellTiter-blue fluorescence Cell Viability Assay (Promega) following the manufacturer’s instructions.
Latent HIV-1 reactivation assay
J-Lat 8.4 and 10.6 clones were used as HIV-1 latency models. J-Lat cell lines are Jurkat cells latently infected with HIV and contain an enhanced green fluorescent protein (GFP) as reporter gene, which is normally expressed at a very low level. When the HIV-1 provirus is reactivated, the protein is expressed at a higher level. Cells were seeded in a 24-well plate at a density of 10⁵ cells per well. Two-fold serial dilutions of JGDM extract (2.5 to 80 µg/mL) were added, followed by incubation at 37°C with 5% CO₂ for 24 hours. Untreated cells (mock) were also incubated and used as a negative control. Moreover, Ingenol-B (0.45 µg/mL), previously identified as LRA by our group[9], was used as positive control. After incubation, the contents of each well were collected and centrifuged at 400 × g for 4 minutes. After centrifugation, the culture medium was removed, the cells were washed with 50 μL of PBS and GFP-positive cells were gated using flow cityometry on BD Accuri C6 cytometer (BD Biosciences). We defined the cells expressing the GFP protein as GFP-positive cells. The percentage of GFP-positive cells represents the level of HIV-1 reactivation. A similar protocol was used for the analysis of PKC inhibitors in J-Lat 10.6 clones. However, in this assay, the cells were pre-incubated for 30 minutes with different PKC inhibitors (GÖ6983, GÖ6976 and RO-31-8220) at 1 μM concentration. Then, the cells were treated with 40 µg/mL of JGDM or Ingenol-B (0.45 µg/mL) for each inhibitor and incubated for 24 hours at 37°C and 5% CO2.
CD4 surface molecules expression evaluation
MT-4 cells were seeded in 24 wells plate at a density of 105 cells per well. Two-fold serial diluted JGDM extract (2.5 to 80 μg/mL) was added, followed by incubation of the plate at 37°C and 5% CO2 for 24 hours. Ingenol-B (0.45 µg/mL) was used as positive control[9]. Untreated cells (mock) were also incubated and used as negative control. After that, the contents of each well were collected and centrifuged at 400 × g for 4 minutes. After centrifugation, culture medium was removed and CD4-Qdot 655 antibody (clone S3.5 - Life Technologies) diluted 10% in 1x PBS was added to each sample. Not labeled untreated cells were used as blank. After 1 hour at -4°C, the CD4 surface marker expression was evaluated gating cells with fluorescence using flow cytometry on BD Accuri C6 cytometer (BD Biosciences). The same protocol used for analysis of PKC inhibitors in J-Lat 10.6 clones was used in MT-4 cells followed by CD4 surface molecules expression evaluation.
Statistical analysis
Cell-based experiments were independently repeated at least three times. CC50 and EC50 were calculated using a non-linear regression (curve fit) in the GraphPad Prism 9.3.0 program. The selectivity index was determined using the formula: SI = CC50/EC50. Statistical analysis was done using Repeated Measures One-Way ANOVA (RM-One Way ANOVA) or Two-Way ANOVA followed by Tukey’s comparison in GraphPad Prism program.
Results and Discussion
JGDM presents antiviral activity against HIV-1
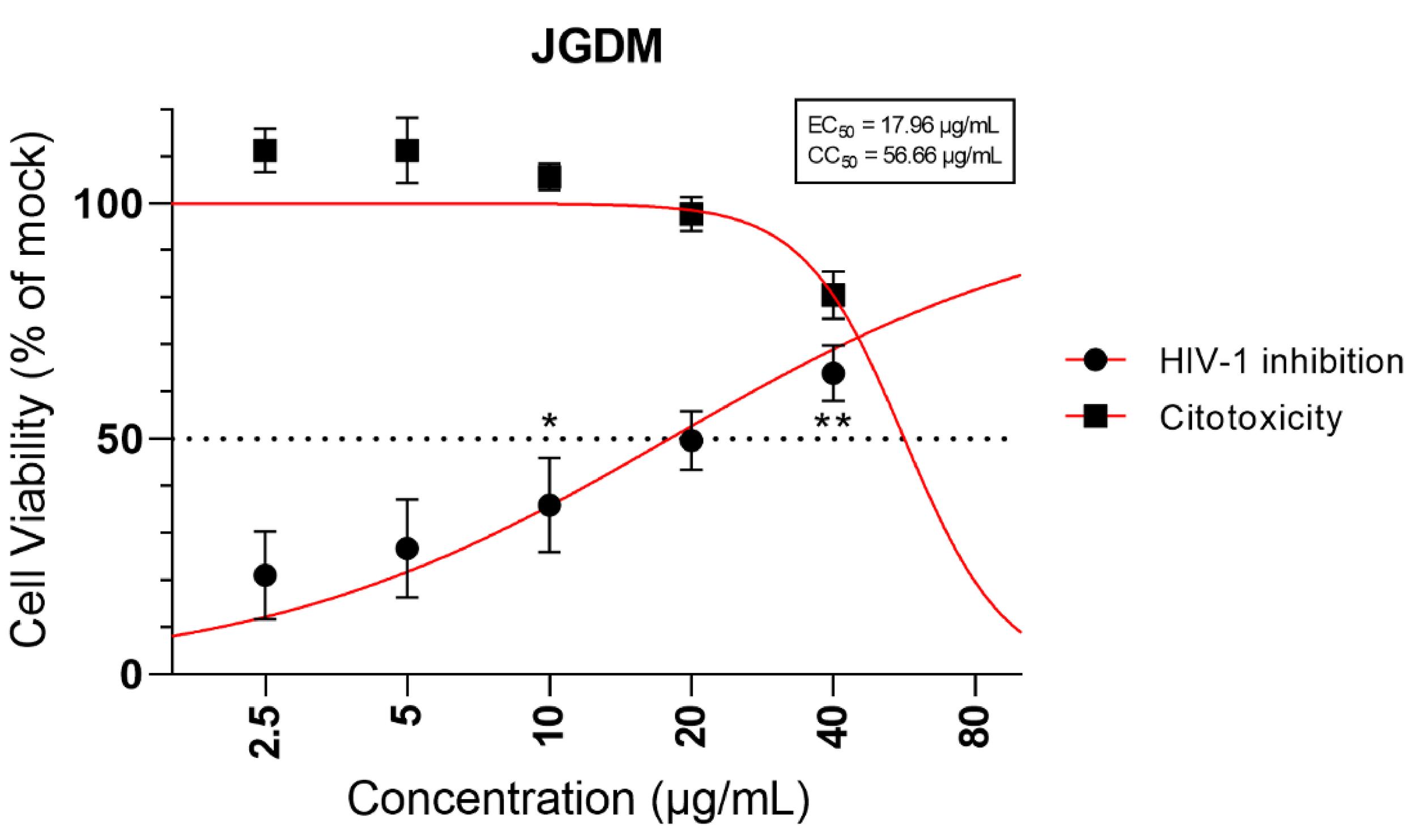
Evaluating the antiviral potential of JGDM, it was observed that JGDM significantly increased (p < 0.05, One-Way ANOVA followed by Tukey’s) the mean of viable cells in a concentration dependent manner from 10 μg/mL to 40 μg/mL with an EC50 of 17.96 μg/mL (FIGURE 1). The 3TC also effectively inhibited HIV-1 infection in MT-4 cells (data not shown), with the effective dose being similar with our tested compound (EC50 = 14.71 μg/mL) and corroborating with previous findings by our group[14].
: Antiviral activity against HIV-1 virus infection and cytotoxicity of different concentrations of the JGDM extract in MT-4 cells. The curve is derived from a non-linear regression whereas the values represented correspond to the mean ± SEM.
* Represents significant difference (p < 0.05, RM-One-Way ANOVA).
Regarding cytotoxicity, JGDM demonstrated low cytotoxic activity at its effective concentration (40 μg/mL), with a CC50 of 56.66 μg/mL (FIGURE 1). Conversely, the 3TC exhibited cytotoxic activity solely above effective concentration against HIV-1, primarily observed between 24-48 μg/mL (data not shown).
For interpretation of the selectivity index results, when SI value is greater than 1, it indicates that the compound has higher selectivity for HIV than for the MT-4 cell, whereas an SI value less than 1 indicates that the compound has a higher toxicity for MT-4 than for HIV. In our study the SI value for JGDM was 3.15. Taken together these data suggest that JGDM has antiviral activity against HIV-1 in MT-4 cells and is more active against HIV than for the cell.
JGDM latent HIV-1 reactivation is mediated by PKC pathway
To test JGDM ability in latency, two HIV-1 latency cell models, J-Lat cells 8.4 and 10.6 clones, were treated with different concentrations of our compound and Ing-B. Our findings indicate that JGDM contains compounds capable of reactivating latent HIV in J-Lat 8.4 cells, with a maximum of 4% of GFP-positive cells at the concentration of 80 μg/mL. In comparison, the Ing-B control enhanced a maximum of 20% GFP-positive cells, representing a 5-fold difference with JGDM (FIGURE 2A and 2C).
: JGDM extract reactivates latent HIV-1 in J-Lat cells 8.4 and 10.6 and occurs through PKC pathway.
Similarly, J-Lat 10.6 cells exhibited higher sensitivity to latent virus reactivation and showed that JGDM extract possesses the ability to reactivate latent HIV-1, significantly increasing (p < 0.01, Two-Way ANOVA followed by Tukey’s comparison) up to 32% of GFP-positive cells in a concentration dependent manner from 10 μg/m to 40 μg/mL (FIGURE 2A and 2C). Although with lower potency compared to Ing-B, it represents a 51% difference with positive control.
Compared to the results obtained with the MT-4 cytotoxicity assay, where 40 μg/mL concentrations showed no cytotoxic activity (FIGURE 1A), we can consider that the sensitivity of J-Lat 8.4 and 10.6 cells to the JGDM at this concentration is lower and had no interference in its activity.
(A) J-Lat cells (8.4 and 10.6 clones) were incubated with either JGDM extract (2.5 to 80 μg/mL) or Ing-B (0.45 μg/mL) for 24 hours and subsequently the expression of GFP-positive cells was detected by flow citometry. Values represented correspond to the mean ± SD of four experiments. ** and **** represent significant difference (p < 0.01 and p < 0.0001, respectively, by Two-Way ANOVA followed by Tukey's comparison). (B) J-Lat cells were pre-treated with different PKC inhibitors (GÖ6976, GÖ6983 and RO-31-8220) at 1 μM concentration for 30 minutes prior to addition of either JGDM extract (40 μg/mL) or Ing -B (0.45 μg/mL). GFP expression was assessed 24 h after treatment by flow cytometry. Values represented correspond to the mean ± SD of five experiments. (C) Flow cytometry histograms of gated J-Lat cells (8.4 and 10.6) showing the percentage of GFP-positive cells when treated either with Ing-B (0.45 μg/mL) or JGDM (40 and 80 μg/mL).
In addition to the reactivation assay in J-Lat 10.6 cells where inhibition by JGDM was more relevant, inhibitors of different PKC isoforms were used to verify the activity of JGDM extract with the inhibition of this pathway.
Our results showed that JGDM extract probably acts through the activation of the PKC isoforms, primarily that regulated by the inhibitor GÖ6983, as evidenced by a significant decrease (p < 0.0001) in the percentage of GFP-positive cells when treated with the 40 μg/mL concentration. Compared to the previous level of approximately 40%, we observed a 30% reduction of HIV-1 reactivation activity (FIGURE 2B)
Similar results were observed with the positive control, suggesting that the active compound present in the JGDM extract likely acts through the same pathway as Ing-B, as evidenced by the substantial reduction in GFP-positive cells when the GÖ6983 inhibitor was present (FIGURE 2B).
JGDM also downregulates CD4 expression through PKC
To investigate the ability of JGDM to change the expression of CD4 molecules on the cell surface, CD4 expression was measured by flow cytometry in MT-4 cells stimulated with JGDM or Ing-B.
Our results showed that JGDM extract has the ability to downregulate the expression of CD4 in human T cell lineage. In comparison with mock, it is observed that JGDM extract significantly reduced (p < 0.0001) up to 66% of the CD4 expression in a concentration dependent manner from 2.5 μg/mL to 40 μg/mL (FIGURE 3A and 3C), meanwhile Ing-B reduced 80% of CD4 expression.
: JGDM downregulates the expression of CD4 molecules in MT-4 cell surface at different concentrations and its downregulatory activity is antagonized by PKC isoforms inhibitors.
To verify the possibility of JGDM extract acting through the PKC isoforms to downregulate CD4 expression, three different inhibitors of PKC isoforms were used. Our results indicate that the downregulation activity of JGDM is associated with the activation and modulation of PKC isoforms. This is evidenced by the fact that, when inhibited by the GÖ6983 inhibitor, JGDM significantly increased (p < 0.0001) CD4 expression by 78%, compared to control CD4 expression of 25% (FIGURE 3B).
Taken together this result shows that the same PKC inhibitor used to inhibit the expression of CD4 molecules on T cell surface also inhibits the latent HIV reactivation by the JGDM extract. Moreover, GÖ6983 inhibitor also reduced the CD4 modulation by Ing-B, reinforcing the hypothesis that JGDM acts through the same pathway as Ing-B and it contains compounds that can be used for the "shock and kill" strategy.
Natural compounds from plants, marine sources, and other products offer potential for new antiviral treatments to enhance current combination antiretroviral therapy (cART) by targeting HIV latent reservoirs and acting as potent inhibitors[15]. However, no clinical study has yet demonstrated consistent reduction of viral reservoirs with single latency-reversing agents (LRAs). This highlights the need for new strategies, including the co-administration of LRAs from different functional classes, as in vitro and ex vivo studies show that combining distinct LRAs can produce synergistic effects greater than when used individually[16-18].
(A) MT-4 cells were stimulated with either different concentrations of JGDM (2.5 to 80 μg/mL) or Ing-B (0.45 μg/mL) for 24 hours. Subsequently, an anti-CD4 antibody was added and expression of the antibody bound to the cell surface CD4 molecules was assessed by flow cytometry. Values represented correspond to the mean ± SD of three experiments. **** represents significant difference (p < 0.0001, Two-Way ANOVA followed by Tukey's comparison). (B) MT-4 cells were pretreated with different PKC inhibitors (GÖ6976, GÖ6983 and RO-31-8220) at 1 μM concentration for 30 minutes prior to the addition of the extract JGDM (40 μg/mL) and Ing-B (0.45 μg/mL) for 24 hours. Subsequently, an anti-CD4 antibody was added and expression of the antibody bound to cell surface CD4 molecules was assessed by flow cytometry. Values represented correspond to the mean ± SD of three experiments. (C) Flow cytometry histograms of CD4 expression in MT-4 cells treated either with JGDM (40 μg/mL) or Ing-B (0.45 μg/mL) showing the mean fluorescence intensity (MFI) decrease when cells are treated.
Futhermore, to be clinically applicable, effective LRAs should be not only highly potent but also display low cytotoxic to immune cells[7]. In summary, our findings revealed that J. gossypiifolia is a source of natural bioactive compounds for LRAs that could be used as support in reactivating latent reservoirs and prevent re-seeding of viral reservoirs, but the main limitation of this study is that the active compound responsible for the HIV reverse activity is not yet know to be clinically used.
Many naturally derived drugs have been found to be safe and have granted FDA approval, and many have progressed to clinical trials[19]. Bryostatin-1, for example, has been tested in over 30 phase I and II clinical trials for a variety of cancers[20]. The cyclic peptide romidepsin, derived from Chromobacterium violaceum, is approved for treating T-cell lymphomas and has also been shown to induce HIV expression. A recent clinical study demonstrated that it increases viral RNA levels in the plasma of aviremic patients[21,22]. Regarding our JGDM extract, it has been shown to be safe and well tolerated in vitro but further testing with the purified active compound would be necessary for it to be applied clinically.
We show here that dichloromethane extract of J. gossypiifolia reactivates latent HIV-1 in vitro and also has antiviral activity against HIV-1. It has been also previously described that other Jatropha genus plant, J. curcas, is able to inhibit HIV-induced cytopathic effects in peripheral blood mononuclear cells (PBMCs) at IC50 = 120 µg/mL[23]. In comparison with JGDM antiviral activity, J. curcas presents a lower potential than JGDM.
Diterpenes from latex or radix of plants from Euphorbiaceae family like Euphorbia lacteal[24], E. tirucalli[9], E. amygdaloides ssp. Semiperfoliata[25] and E. kansui[26] have been studied to reactivate HIV-1 provirus in vitro and to inhibit HIV-1 infection. In comparison with our reactivation positive control, Ing-B, previously isolated from E. tirucalli, JGDM is less effective in reactivating latent HIV-1 (50% of GFP+ cells with 0.01 µM) and inhibiting HIV-1 infection (EC50 =0.022 µM)[9]. This could be explained by other compounds present in the dichloromethane fraction that can be interfering in these activities.
Therefore, we investigated the role of the PKC pathway in reactivating latent HIV-1. Our findings indicate that JGDM's reactivation activity is reduced by the pan-PKC inhibitor GÖ6983 and likely functions as a PKC agonist.
PKCs belong to the superfamily of serine/threonine kinases whose members are differentially regulated by intracellular signals transduction such as increase in calcium concentration, diglycerides and phosphatidic acid[27]. There are at least ten PKC isoforms which can be subdivided into three subfamilies based on their domain structure and cofactor requirement. The classical or conventional PKC isoforms (α, βI, βII and γ) can be regulated by phospholipids and diacylglycerol (DAG) as well as Ca2+; novel PKC isoforms (δ, ε, θ, η and μ) can be regulated by phospholipids and DAG but not by Ca2+; while atypical isoforms (ζ and ι/λ) do not respond to either DAG or Ca2+, but can be regulated by phospholipids[27,28].
Regarding the PKC inhibitors specificity, studies shows that GÖ6983 inhibits atypical isoform PKCζ besides classical (PKCα, β and γ) and new isoforms (only δ)[29]. RO-31-8220, on the other hand, has the ability to inhibit the isoforms PKCα, PKCβI, PKCβII, PKCγ and PKCε[30]. Considering that the atypical isoforms are not dependent on calcium ions and DAG for its activation, is very unlikely that JGDM has an impact in this isoform. Moreover, Hezareh et al.[29] found that compounds like diterpenes do not act through this isoform.
Previously studies reported other PKC agonists isolated from Euphorbiaceae family plants, such as prostratin[31], 4β-dPE A[25], PEP005[8], EK-15A[26] or SJ23B[32]. Other natural compounds like bryostatin-1 have been also used to reactivate HIV in different cell models and as a partial agonist of PKC[33]. Same way as the JGDM reported here, bryostatin-1 can reactivates HIV-1 through a classical PKC-dependent pathway (reduced by the PKC inhibitor GÖ6983) and, as expected, downregulates the expression of the HIV-1 co-receptors CD4 and CXCR4 preventing de novo HIV-1 infection in susceptible cells[17].
We further investigated whether the anti-HIV activity was partly due to CD4 downregulation. Our data showed that JGDM effectively downregulates CD4 surface expression, and this effect is reduced by the PKC inhibitor GÖ6983. HIV entry involves the interaction of gp120 and gp41 with CD4, CXCR4 and/or CCR5 receptors[34]. It has been reported that the anti-HIV activity of prostratin is probably due to induction of CD4 and CXCR4 downregulation by promoting endocytosis of CD4 receptors and degradation of these molecules[29]. This internalization can occur because PKC phosphorylation of specific serine residues in the cytoplasmic domain of the molecule and dissociation of p56lck leads to CD4 recruitment into clathrin-coated pits and its internalization[35]. Therefore, the decreased surface expression of CD4 observed with JGDM treatment may be an important factor in the blockade of de novo infection and a possible mechanism of action involved in antiviral activity since as our results showed decreased levels of CD4 molecules in JGDM treated cells, the virus is unable to interact with entry receptor and consequently to infect the cells.
In addition, when cells were treated with PKC inhibitors, CD4 expression returned to normal levels. Previous studies showed that compounds like diterpenoids that modulates the PKC pathway, presents a common pathway-based mechanism of action for inactivation of both HIV and Chikungunya virus (CHIKV)[36]. Indeed, our group has found that the bioactive compound present in JGDM extract responsible for the PKC-dependent latent reactivation, CD4 downregulation and inhibition of HIV-1 infection also presents inhibitory activity for the CHIKV infection on Vero cells (data not shown).
Conclusion
Therefore, here in this work we show that JGDM extract has antiviral activity against HIV-1 and can perform the reactivation of latent HIV-1 in J-Lat different cell lines, but with a lower potential than other compounds. In addition, both reactivation and CD4 downmodulation performed by JGDM are modulated by PKC isoforms and probably acts through the classical and/or new PKC isoforms, as well as the previously reported PKC agonist Ingenol-B. These findings are encouraging and further safety and efficacy in vitro and in vivo studies will be performed to identify the chemical constituents of this extract and to better define the mechanism of action.
Acknowledgments
We are grateful for the financial support provided by the Federal Government Agency CAPES.
References
- 1 Antiretroviral Therapy Cohort C. Survival of HIV-positive patients starting antiretroviral therapy between 1996 and 2013: a collaborative analysis of cohort studies. Lancet HIV. 2017; 4(8): e349-e356. [ https://doi.org/10.1016/S2352-3018 (17)30066-8] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc5555438/ ].
» https://doi.org/10.1016/S2352-3018 (17)30066-8» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc5555438/ - 2 Brady MT, Oleske JM, Williams PL, Elgie C, Mofenson LM, Dankner WM, et al. Declines in mortality rates and changes in causes of death in HIV-1 infected children during the HAART era. J Acquir Immune Defic Syndr. 2010; 53(1): 86-94. [ https://doi.org/10.1097/qai.0b013e3181b9869f ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc2801894/ ].
» https://doi.org/10.1097/qai.0b013e3181b9869f» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc2801894/ - 3 Murray AJ, Kwon KJ, Farber DL, Siliciano RF. The latent reservoir for HIV-1: How immunologic memory and clonal expansion contribute to HIV-1 persistence. J Immunol. 2016; 197(2): 407-17. [ https://doi.org/10.4049/jimmunol.1600343 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4936486/ ].
» https://doi.org/10.4049/jimmunol.1600343» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4936486/ - 4 Archin NM, Margolis DM. Emerging strategies to deplete the HIV reservoir. Curr Opin Infect Dis. 2014; 27(1): 29-35. [ https://doi.org/10.1097%2FQCO.0000000000000026 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4031321/ ].
» https://doi.org/10.1097%2FQCO.0000000000000026» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4031321/ - 5 Deeks SG. HIV: shock and kill. Nature. 2012; 487(7408): 439-440. [ https://doi.org/10.1038/487439a ].
» https://doi.org/10.1038/487439a - 6 Rasmussen TA, Tolstrup M, Sogaard OS. Reversal of latency as part of a cure for HIV-1. Trends Microbiol. 2016; 24: 90-97. [ https://doi.org/10.1016/j.tim.2015.11.003 ].
» https://doi.org/10.1016/j.tim.2015.11.003 - 7 Jiang G, Dandekar S. Targeting NF-kappaB signaling with protein kinase C agonists as an emerging strategy for combating HIV latency. AIDS Res Hum Retrovir. 2015; 31(1): 4-12. [ https://doi.org/10.1089/aid.2014.0199 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4287114/ ].
» https://doi.org/10.1089/aid.2014.0199» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4287114/ - 8 Jiang G , Mendes EA , Kaiser P , Wong DP , Tang Y , Cai I , et al . Synergistic reactivation of latent HIV expression by ingenol-3-angelate, PEP005, targeted NF-?B signaling in combination with JQ1 induced p-TEFb activation. PLoS Pathog. 2015; 11(7): e1005066. [ https://doi.org/10.1371/journal.ppat.1005066 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4520526/ ].
» https://doi.org/10.1371/journal.ppat.1005066» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4520526/ - 9 Abreu CM, Price SL, Shirk EN, Cunha RD, Pianowski LF, Clements JE, et al. Dual role of novel ingenol derivatives from Euphorbia tirucalli in HIV replication: Inhibition of de novo infection and activation of viral LTR. PLoS One. 2014; 9(5): 1-14. [ https://doi.org/10.1371/journal.pone.0097257 ]. [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4020785/ ].
» https://doi.org/10.1371/journal.pone.0097257» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4020785/ - 10 Mehla R, Bivalkar-Mehla S, Zhang R, Handy I, Albrecht H, Giri S, et al. Bryostatin modulates latent HIV-1 infection via PKC and AMPK signaling but inhibits acute infection in a receptor independent manner. Plos One. 2010; 5(6): 1-15. [ https://doi.org/10.1371/journal.pone.0011160 ]. [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc2886842/ ].
» https://doi.org/10.1371/journal.pone.0011160» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc2886842/ - 11 Zhao M , De Crignis E , Rokx C , Verbon A , van Gelder T , Mahmoudi T , et al . T cell toxicity of HIV latency reversing agents. Pharmacol Res. 2019; 139: 524-534. [ https://doi.org/10.1016/j.phrs.2018.10.023 ].
» https://doi.org/10.1016/j.phrs.2018.10.023 - 12 Félix-Silva J, Giordani RB, Silva-Jr AA, Zucolotto SM, Fernandes-Pedrosa MF. Jatropha gossypiifolia L. (Euphorbiaceae): A review of traditional uses, phytochemistry, pharmacology, and toxicology of this medicinal plant. Evid Based Complement Alternat Med. 2014; 2014(369204): 1-32. [ https://doi.org/10.1155/2014/369204 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4070477/ ].
» https://doi.org/10.1155/2014/369204» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4070477/ - 13 Falodun A, Kragl U, Touem SM, Villinger A, Fahrenwaldt T, Langer P. A novel anticancer diterpenoid from Jatropha gossypifolia Nat Prod Commun. 2012; 7(2): 151-152. [ https://doi.org/10.1177%2F1934578X1200700204 ].
» https://doi.org/10.1177%2F1934578X1200700204 - 14 Valadão ALC , Abreu CM , Dias JZ , Arantes P , Verli H , Tanuri A , et al . Natural plant alkaloid (Emetine) inhibits HIV-1 replication by interfering with reverse transcriptase activity. Molecules. 2015; 20(6): 11474-11489. [ https://doi.org/10.3390/molecules200611474 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc6272240/ ].
» https://doi.org/10.3390/molecules200611474» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc6272240/ - 15 Andersen RJ , Ntie-Kang F and Tietjen I . Natural product-derived compounds in HIV suppression, remission, and eradication strategies. Antiviral Res. 2018; 158: 63-77. [ https://doi.org/10.1016/j.antiviral.2018.07.016 ].
» https://doi.org/10.1016/j.antiviral.2018.07.016 - 16 Reuse S, Calao M, Kabeya K, Guiguen A, Gatot JS, Quivy V, et al. Synergistic activation of HIV-1 expression by deacetylase inhibitors and prostratin: Implications for treatment of latent infection. Plos One. 2009; 4(6): e6093. [ https://doi.org/10.1371/journal.pone.0006093 ] [ https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2699633/ ].
» https://doi.org/10.1371/journal.pone.0006093» https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2699633/ - 17 Pérez M, Vinuesa AG, Sanchez-Duffhues G, Marquez N, Bellido ML, Muñoz-Fernandez A, et al. Bryostatin-1 synergizes with histone deacetylase inhibitors to reactivate HIV-1 from latency. Curr HIV Res. 2010; 8(6): 418-429. [ https://doi.org/10.2174/157016210793499312 ].
» https://doi.org/10.2174/157016210793499312 - 18 Laird GM, Bullen CK, Rosenbloom DIS, Martin AR, Hill AL, Durand CM, et al. Ex vivo analysis identifies effective HIV-1 latency-reversing drug combinations. J Clin Invest. 2015; 125(5): 1901-1912. [ https://doi.org/10.1172%2FJCI80142 ] [ https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4463209/ ].
» https://doi.org/10.1172%2FJCI80142» https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4463209/ - 19 Banerjee S , Nau S , Hochwald SN , Xie H , Zhang J . Anticancer properties and mechanisms of botanical derivatives. Phytomed Plus. 2023; 3(1): 100396. [ https://doi.org/10.1016/j.phyplu.2022.100396 ].
» https://doi.org/10.1016/j.phyplu.2022.100396 - 20 Kollár P, Rajchard J, Balounová Z, Pazourek J. Marine natural products: Bryostatins in preclinical and clinical studies. Pharm Biol. 2014; 52(2): 237-242. [ https://doi.org/10.3109/13880209.2013.804100 ].
» https://doi.org/10.3109/13880209.2013.804100 - 21 Bertino EM , Otterson GA . Romidepsin: A novel histone deacetylase inhibitor for cancer. Expert Opin Investig Drugs. 2011; 20(8): 1151-1158. [ https://doi.org/10.1517/13543784.2011.594437 ].
» https://doi.org/10.1517/13543784.2011.594437 - 22 Søgaard OS, Graversen ME, Leth S, Olesen R, Brinkmann CR, Nisse SK, et al. The depsipeptide romidepsin reverses HIV-1 latency in vivo. Plos Pathog. 2015; 11(9): e1005142. [ https://doi.org/10.1371/journal.ppat.1005142 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4575032/ ].
» https://doi.org/10.1371/journal.ppat.1005142» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4575032/ - 23 Dahake R , Roy S , Patil D , Rajopadhye S , Chowdhary A , Deshmukh RA . Potential anti-HIV activity of Jatropha curcas Linn. leaf extracts. J Antivir Antiretrovir. 2013; 5: 7. [ https://doi.org/10.4172/jaa.1000082 ].
» https://doi.org/10.4172/jaa.1000082 - 24 Avila L, Perez M, Sanchez-Duffhues G, Hernandez-Galan R, Munoz E, Cabezas F, et al. Effects of diterpenes from latex of Euphorbia lactea and Euphorbia laurifolia on human immunodeficiency virus type 1 reactivation. Phytochemistry. 2010; 71(2-3): 243-248. [ https://doi.org/10.1016/j.phytochem.2009.10.005 ].
» https://doi.org/10.1016/j.phytochem.2009.10.005 - 25 De la Torre-Tarazona HE , Jiménez R , Bueno P , Camarero S , Román L , Fernández-García JL , et al . 4-Deoxyphorbol inhibits HIV-1 infection in synergism with antiretroviral drugs and reactivates viral reservoirs through PKC/MEK activation synergizing with vorinostat. Biochem Pharmacol. 2020; 177: 113937. [ https://doi.org/10.1016/j.bcp.2020.113937 ].
» https://doi.org/10.1016/j.bcp.2020.113937 - 26 Yang H , Li X , Yang X , Lu P , Wang Y , Jiang Z , et al . Dual effects of the novel ingenol derivatives on the acute and latent HIV-1 infections. Antiviral Res. 2019; 169: 104555. [ https://doi.org/10.1016/j.antiviral.2019.104555 ].
» https://doi.org/10.1016/j.antiviral.2019.104555 - 27 Kazi JU, Kabir NN, Rönnstrand L. Protein kinase C (PKC) as a drug target in chronic lymphocytic leukemia. Med Oncol. 2013; 30(4): 757. [ https://doi.org/10.1007/s12032-013-0757-7 ].
» https://doi.org/10.1007/s12032-013-0757-7 - 28 Wu-Zhang AX, Newton AC. Protein kinase C pharmacology: refining the toolbox. Biochem J. 2013; 452(2): 195-209. [ https://doi.org/10.1042/bj20130220 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4079666/ ].
» https://doi.org/10.1042/bj20130220» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4079666/ - 29 Hezareh M , Moukil MA , Szanto I , Pondarzewski M , Mouche S , Cherix N , et al . Mechanisms of HIV receptor and co-receptor down-regulation by prostratin: role of conventional and novel PKC isoforms. Antivir Chem Chemother. 2004; 15: 207-222. [ https://doi.org/10.1177/095632020401500404 ].
» https://doi.org/10.1177/095632020401500404 - 30 Harris TE, Persaud SJ, Jones PM. Atypical isoforms of pKc and insulin secretion from pancreatic beta-cells: evidence using Gö6976 and Ro31-8220 as PKC inhibitors. Biochem Biophys Res Commun. 1996; 227(3): 672-676. [ https://doi.org/10.1006/bbrc.1996.1567 ].
» https://doi.org/10.1006/bbrc.1996.1567 - 31 Biancotto A, Grivel JC, Gondois-Rey F, Bettendroffer L, Vigne R, Brown S, et al. Dual role of prostratin in inhibition of infection and reactivation of human immunodeficiency vírus from latency in primary blood lymphocytes and lymphoid tissue. J Virol. 2004; 78(19): 10507-10515. [ https://doi.org/10.1128%2FJVI.78.19.10507-10515.2004 ] [ https://pubmed.ncbi.nlm.nih.gov/15367617 ].
» https://doi.org/10.1128%2FJVI.78.19.10507-10515.2004» https://pubmed.ncbi.nlm.nih.gov/15367617 - 32 Bedoya LM , Márquez N , Martínez N , Gutiérrez-Eisman S , Álvarez A , Calzado MA , et al . SJ23B, a jatrophane diterpene activates classical PKCs and displays strong activity against HIV in vitro. Biochem Pharmacol. 2009; 77: 965-978. [ https://doi.org/10.1016/j.bcp.2008.11.025 ].
» https://doi.org/10.1016/j.bcp.2008.11.025 - 33 Díaz L , Martínez-Bonet M , Sánchez J , Fernández-Pineda A , Jiménez JL , Muñoz E , et al . Bryostatin activates HIV-1 latent expression in human astrocytes through a PKC and NF-?B-dependent mechanism. Sci Rep. 2015; 5: 12442. [ https://doi.org/10.1038/srep12442 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4510492/ ].
» https://doi.org/10.1038/srep12442» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc4510492/ - 34 Klasse PJ. The molecular basis of HIV entry. Cell Microbiol. 2012; 14(8): 1183-1192. [ https://doi.org/10.1111/j.1462-5822.2012.01812.x ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc3417324/ ].
» https://doi.org/10.1111/j.1462-5822.2012.01812.x» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc3417324/ - 35 Pelchen-Matthews A, Parsons IJ, Marsh M. Phorbol ester-induced downregulation of CD4 is a multistep process involving dissociation from p56lck, increased association with clathrin-coated pits, and altered endosomal sorting. J Exp Med. 1993; 178(4): 1209-1222. [ https://doi.org/10.1084/jem.178.4.1209 ] [ http://www.ncbi.nlm.nih.gov/pmc/articles/pmc2191214/ ].
» https://doi.org/10.1084/jem.178.4.1209» http://www.ncbi.nlm.nih.gov/pmc/articles/pmc2191214/ - 36 Nothias-Scaglia LF, Pannecouque C, Renucci F, Delang L, Neyts J, Roussi F, et al. Antiviral activity of diterpene esters on Chikungunya virus and HIV replication. J Nat Prod. 2015; 78(6): 1277-1283. [ https://doi.org/10.1021/acs.jnatprod.5b00073 ].
» https://doi.org/10.1021/acs.jnatprod.5b00073
-
Funding:
This research received no external funding.
Publication Dates
- Publication in this collection
10 July 2025 - Date of issue
June 2025
History
- Received
29 July 2024 - Accepted
03 June 2025

 Source: Authors, 2024.
Source: Authors, 2024.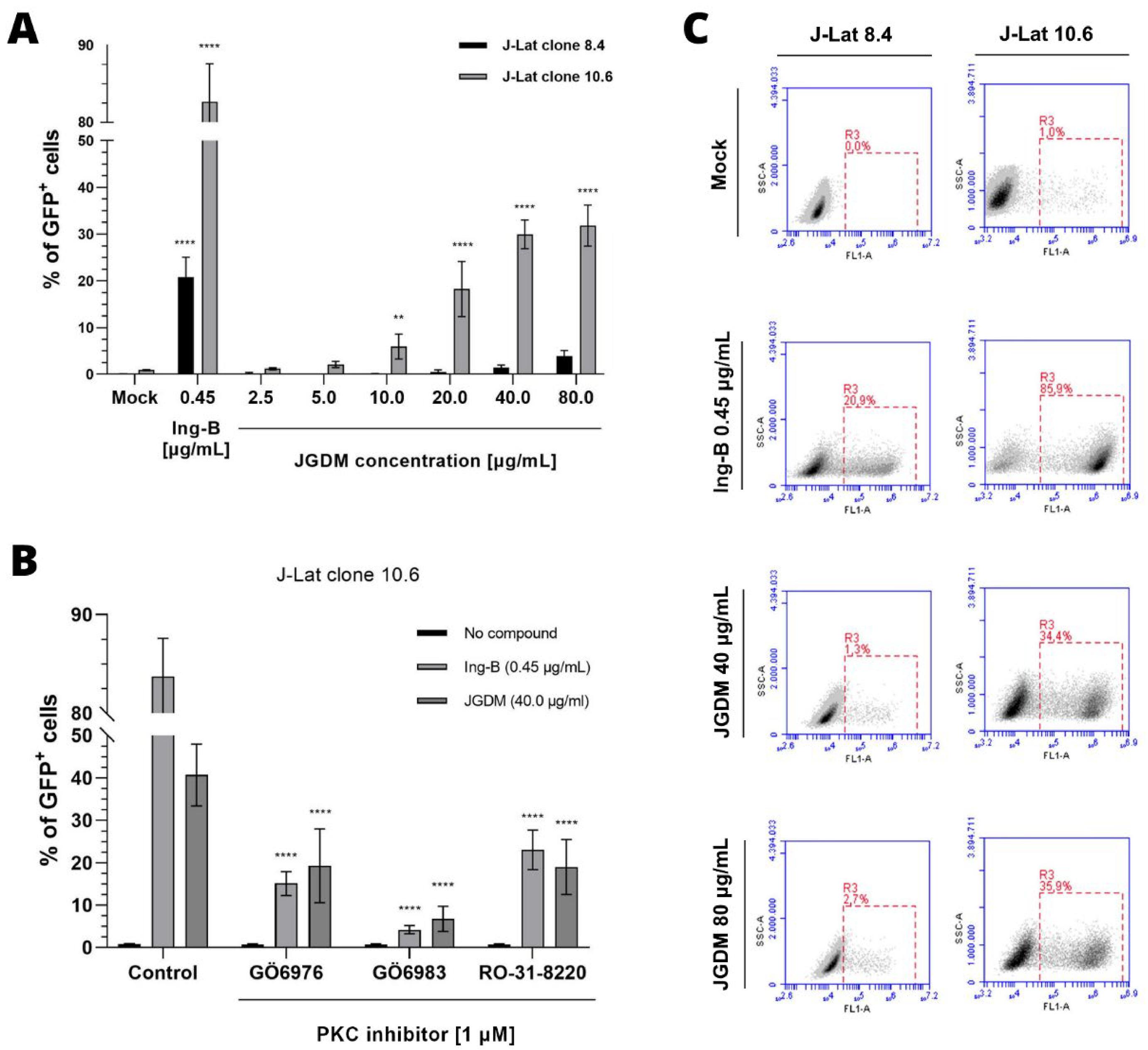 Source: Authors, 2024.
Source: Authors, 2024.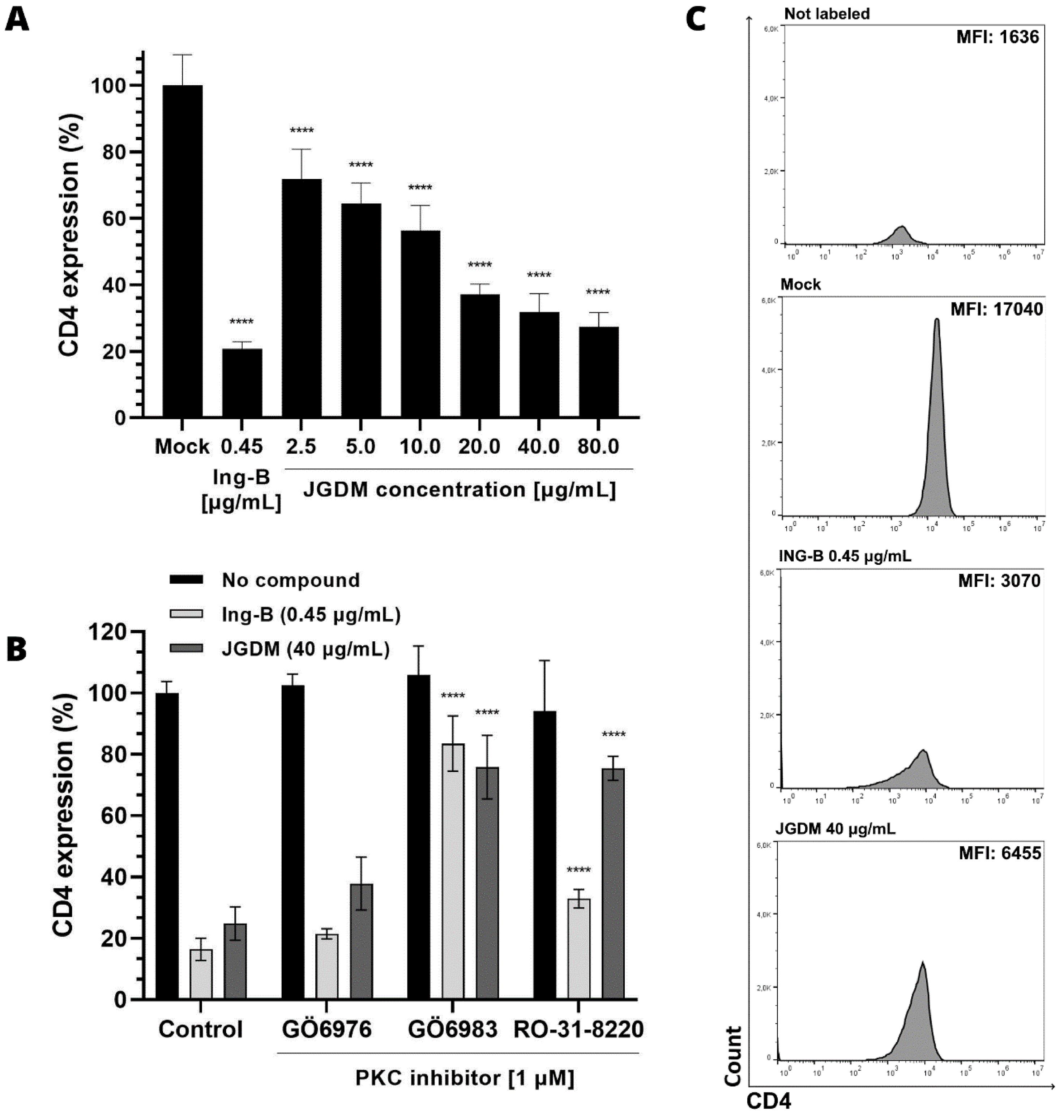 Source: Authors, 2024.
Source: Authors, 2024.