Resumo
O canabidiol (CBD) é um fitocanabinoide que apresenta várias propriedades psicotrópicas com potencial terapêutico que têm sido melhor compreendidas através de estudos pré-clínicos que avaliam os alvos moleculares e regiões encefálicas envolvidas. Esta revisão narrativa compila e discute estudos que investigaram os efeitos do CBD sobre a extinção e reconsolidação de memórias aversivas, bem como sobre a expressão de medo e ansiedade em ratos e camundongos. Os resultados obtidos sugerem que a administração de CBD prejudica a reconsolidação de memórias aversivas, pode acelerar a extinção, além de reduzir comportamentos relacionados ao medo e à ansiedade. Essas ações envolvem principalmente a ativação de receptores canabinoides do tipo 1 (CB1) e de serotonina do tipo 1A (5-HT1A) expressos em diferentes regiões encefálicas. Entretanto, os efeitos do CBD em protocolos que induzem a formação de memórias com características traumáticas e respostas de medo/ansiedade inadequadas ainda permanecem poucos explorados. Outras três lacunas de conhecimento a serem exploradas incluem as diferenças sexuais, a suscetibilidade à interferência em memórias aversivas/traumáticas remotas, e os efeitos de regimes de tratamento repetidos. Compreender as bases farmacológicas do CBD é essencial para estabelecer sua eficácia, segurança e potencial aplicação no tratamento de transtornos de ansiedade e estresse em humanos.
Cannabis; Córtex pré-frontal; Amígdala; Hipocampo; THC
Abstract
Cannabidiol (CBD) is a phytocannabinoid with various psychotropic properties and therapeutic potential, as demonstrated by preclinical studies that have identified its molecular targets, and the brain regions involved in its effects. In this narrative review, we compiled and discussed studies that investigated the effects of CBD on the extinction and reconsolidation of aversive memories and the expression of fear and anxiety in rats and mice. Their results suggest that CBD administration impairs the reconsolidation of aversive memories, may accelerate their extinction, and reduce fear- and anxiety-related behaviors. These actions primarily involve the activation of cannabinoid type 1 (CB1) receptor and serotonin type 1A (5-HT1A) receptor expressed in different brain regions. However, the effects of CBD in protocols that induce memories with traumatic characteristics and inappropriate fear/anxiety responses remain underexplored. Three other knowledge gaps to be explored include sexual differences, susceptibility to interference in remote aversive/traumatic memories, and the effects of repeated treatment regimens. Understanding the pharmacological bases of CBD is essential for establishing its efficacy, safety, and potential application in the treatment of anxiety and stress disorders in humans.
Cannabis; Medial prefrontal córtex; Amygdala; hippocampus; THC
Introdução
O canabidiol (CBD), principal fitocanabinoide não psicotomimético da Cannabis sativa, apresenta propriedades farmacológicas com potencial aplicação terapêutica[1] e de relevância para pacientes com transtornos de ansiedade e estresse[2]. Dentre elas, destacam-se a capacidade de atenuar a expressão do medo/ansiedade e modular memórias aversivas via facilitação da extinção e/ou prejuízo da reconsolidação[3-5]. Os mecanismos de ação através dos quais o CBD promove tais efeitos são complexos e envolvem os sistemas de neurotransmissão endocanabinoide e serotoninérgico e vários receptores, tais como os para canabinoides dos tipos 1 e 2 (CB1 e CB2) e para serotonina do tipo 1A (5-HT1A), que estão expressos em áreas encefálicas que modulam aspectos mnemônicos e emocionais[3,4]. A FIGURA 1 apresenta a linha do tempo dos estudos pioneiros que investigaram os efeitos do CBD sobre a ansiedade, extinção e reconsolidação de memórias aversivas em animais de laboratório. A maioria desses estudos foi conduzida no Brasil, inclui o Professor Francisco Silveira Guimarães e/ou seus egressos entre os autores, e foi realizado nos Departamentos de Farmacologia da Universidade de São Paulo (Ribeirão Preto, SP), da Universidade Federal de Santa Catarina (Florianópolis, SC) e da Universidade Federal do Paraná (Curitiba, PR).
: Timeline da publicação dos artigos científicos pioneiros que avaliaram os efeitos do canabidiol (CBD) sobre a ansiedade, extinção e reconsolidação de memórias aversivas em animais de laboratório
Testes para medir o medo e a ansiedade em animais de laboratório (FIGURA 2)
Em estudos pré-clínicos com roedores (ratos e camundongos), a avaliação de comportamentos defensivos relacionados ao medo e ansiedade é feita por meio de uma série de testes que, também, desencadeiam respostas autonômicas e endócrinas expressas quando eles são expostos a ameaças reais ou potenciais[6]. Esses testes permitem avaliar tanto respostas defensivas inatas quanto aprendidas/adquiridas através da experiência e da interação com o ambiente. Eles também permitem avaliar o efeito ansiolítico ou ansiogênico de uma intervenção farmacológica, como no caso da administração única ou repetida de CBD tanto por via sistêmica como diretamente em alguma área encefálica específica de interesse.
Embora o medo e a ansiedade estejam relacionados e compartilhem mecanismos neurobiológicos, eles costumam ser diferenciados em animais de laboratório. Comportamentos relacionados à ansiedade, como a avaliação de risco, geralmente surgem ao se aproximar de ameaças potenciais, mas que ainda estão distantes. Já as respostas de medo, como congelamento e fuga, seriam desencadeadas por ameaças imediatas que já foram identificadas[7]. Ressalta-se, também, que o medo ou ansiedade e a atividade exploratória geralmente estão inversamente relacionadas: a indução de medo ou ansiedade tende a diminuir a atividade exploratória, enquanto a administração de drogas ansiolíticas aumenta esse parâmetro.
Para avaliar comportamentos relacionados à ansiedade, ratos e camundongos podem ser expostos ao teste do labirinto em cruz elevado (FIG. 2A), onde o tempo dispendido nos braços abertos e fechados é medido. Visto que ambientes abertos e iluminados são naturalmente considerados mais aversivos pelos redores devido à maior exposição e mais fácil detecção por um eventual predador, drogas ansiolíticas aumentam o tempo nos braços abertos sem alterar o número de entradas nos braços fechados, que refletiria a atividade exploratória/locomotora do animal[8]. De forma semelhante, o teste da caixa clara/escura (FIG. 2B)9 pode ser utilizado para avaliar comportamentos relacionados à ansiedade, onde espera-se que drogas ansiolíticas aumentem o tempo e/ou número de entradas do animal no compartimento claro do aparato. No teste de interação social (FIG. 2C)10, a redução do medo/ansiedade tende a aumentar a atividade exploratória dos animais. Já nos testes do beber punido de Vogel[11] (FIG. 2D) e da hipofagia induzida pela novidade (novelty-suppressed feeding)12(FIG. 2E), drogas ansiolíticas aumentam a ingestão hídrica e alimentar (ou reduzem o tempo para iniciar tal resposta), mesmo na presença de estímulos aversivos, tais como a aplicação de choques elétricos ou exposição a ambientes novos e/ou iluminados. O teste de esconder esferas (marble-burying test) (FIG. 2F) também é potencialmente relevante para se estudar comportamentos ansiosos em roedores, que são atenuados por drogas utilizadas na clínica para o tratamento do transtorno obsessivo compulsivo[13]. Por fim, o teste do campo aberto (FIG. 2G) nos permite avaliar a ansiedade e a locomoção[14]. Para isso, é medido o tempo que o animal explora o centro e a periferia da arena, respectivamente, sendo observado um aumento no primeiro parâmetro após a administração de drogas ansiolíticas.
: Testes usados para avaliar comportamentos relacionados à ansiedade e/ou medo em animais de laboratório após a administração de canabidiol nos estudos revisados.
Quanto à avaliação de respostas comportamentais relacionados ao medo, no caso dos testes de exposição ao predador (ex: gato) ou ao seu odor (FIG. 2H), a resposta de congelamento, esquiva ou avaliação de risco expressa pelo roedor é mensurada[15]. Em outro teste, o do labirinto em T elevado[16] (FIG. 2I), se quantifica o tempo que o animal leva para sair do braço fechado do aparato em três exposições consecutivas (esquiva inibitória) e o tempo que ele leva para escapar de um dos braços abertos (fuga), comportamentos que são atenuados por drogas ansiolíticas e panicolíticas, respectivamente. Já a estimulação elétrica ou química da substância cinzenta periaquedutal (FIG. 2J) em roedores mimetizaria o que seria equivalente aos ataques de pânico em humanos[17]. Por fim, existem vários protocolos de estresse agudo e crônico que podem ser usados para induzir ou potencializar respostas defensivas relacionados ao medo e ansiedade em roedores, tais como o estresse de contenção e o estresse crônico imprevisível[6].
Protocolos para induzir memórias aversivas e avaliar a extinção e reconsolidação delas em animais de laboratório (FIGURA 3)
A expressão do medo que requer experiência prévia ou treinamento pode ser medida pela observação do comportamento de congelamento (freezing) nos protocolos de condicionamento aversivo clássico ou Pavloviano. Nesses protocolos, roedores aprendem a associar um estímulo neutro a um estímulo aversivo (estímulo incondicionado, EI). Após pareamentos repetidos, o estímulo que antes era neutro se torna um estímulo condicionado (EC), visto que o animal passa a expressar o comportamento de congelamento mesmo na ausência do EI. O estímulo neutro a ser condicionado ao choque elétrico (EI) pode ser um contexto (a própria caixa de condicionamento; FIG. 3A), som, luz ou odor.
: Esquema de protocolos experimentais utilizados nos estudos revisados para induzir a formação de memórias aversivas e avaliar os efeitos do canabidiol (CBD) sobre a extinção e reconsolidação em animais de laboratório.
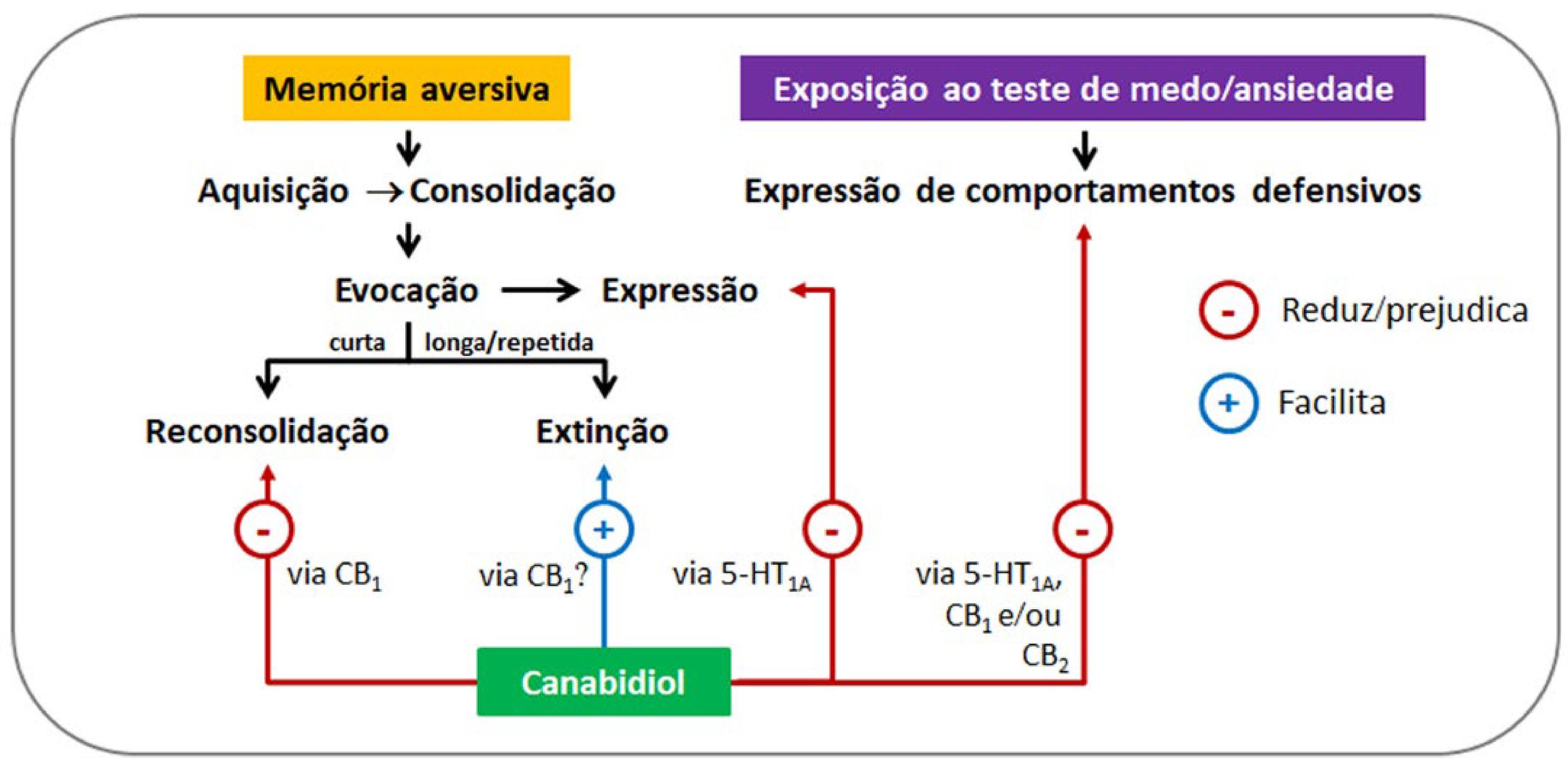
Legenda: (A) No condicionamento aversivo contextual, um ambiente neutro é pareado com choque(s). No teste, o animal expressa congelamento (freezing) quando é recolocado nesse ambiente que se tornou aversivo porque ele lembra da experiencia prévia que teve lá. Uma reexposição curta ao contexto pareado favorece a reconsolidação dessa memória, já uma reexposição mais longa favorece a formação de uma memória de extinção, que compete com a memória aversiva original e a suprime comportamentalmente. Nos casos da esquiva ativa (B) e da esquiva passiva (C), a lógica é similar a supramencionada, mas o comportamento mensurado é a esquiva (avoidance).
Além do comportamento de congelamento, a expressão do medo aprendido em roedores pode ser avaliada pela latência para entrar em um compartimento aversivo (ou o tempo despendido nele) em protocolos baseados em esquiva, também utilizados para a formação de memórias aversivas. Na tarefa de esquiva ativa (FIG. 3B), o animal é colocado em uma câmara com dois compartimentos, onde um choque elétrico é administrado em suas patas em um dos compartimentos, sendo que o roedor aprende a se mover para o outro compartimento após um sinal específico (ex.: um som), a fim de evitar o choque. Em contraste, nas tarefas de esquiva passiva, ao invés de se deslocar para escapar do estímulo aversivo, o animal aprende a permanecer sobre uma plataforma (FIG. 3C) ou em um compartimento específico, evitando assim o recebimento do choque nas patas[18].
A extinção e a reconsolidação são processos dissociáveis que podem ocorrer após a evocação de memórias aversivas formadas em protocolos de condicionamento ou esquiva[19]. Em roedores, a extinção reduz comportamentos defensivos relacionados ao medo, tais como o congelamento e esquiva, por meio da apresentação longa/repetida do EC (um contexto, som, luz ou odor) na ausência do EI (choque). Durante a extinção, uma memória neutra é formada e suprime a memória original, mas não a apaga[20]. Já a reconsolidação acontece após a desestabilização de uma memória, o que a torna suscetível a modificação/atualização até que ela seja reestabilizada novamente[21). Certas intervenções comportamentais e farmacológicas são capazes de interferir com a taxa de extinção e reconsolidação/reestabilização de memórias aversivas, o que pode atenuar seus desfechos comportamentais de forma duradoura [4,22].
Devido às limitações dos biomarcadores atuais para os transtornos de ansiedade e estresse[23,24], estudos comportamentais em animais são essenciais para elucidar as bases neurobiológicas dessas condições psiquiátricas e guiar o desenvolvimento de abordagens terapêuticas mais eficazes. Esta revisão narrativa resume e discute estudos que investigaram os efeitos do CBD na extinção e reconsolidação de memórias aversivas, bem como na expressão do medo aprendido e da ansiedade em ratos e camundongos. Os efeitos do fitocanabinoide delta-9-tetraidrocanabinol (D9-THC) administrado sozinho ou em combinação com o CBD sobre os temas supracitados já foram revisados em outras trabalhos[3,4,25] e, assim, foram excluídos deste estudo.
Metodologia
Este trabalho consistiu em uma revisão narrativa, elaborada de acordo com as diretrizes da Scale for the Assessment of Narrative Review Articles (SANRA). A busca pelos estudos científicos que investigaram os efeitos do CBD sobre comportamentos defensivos relacionados à ansiedade/medo e os processos de extinção e reconsolidação de memórias aversivas em roedores foi conduzida por meio da base de dados PubMed. Para isso, foram utilizados descritores combinados por operadores booleanos, incluindo: *("cannabidiol" OR "CBD") AND ("fear" OR "anxiety" OR "aversive memory" OR “fear memory”) AND ("extinction" OR "reconsolidation") AND ("rodents" OR "rats" OR "mice")*. A busca foi realizada até janeiro de 2025, sem restrições quanto ao ano de publicação, mas limitada a estudos em inglês. A triagem inicial foi baseada na leitura dos títulos e resumos, com exclusão de trabalhos de revisão, que não envolviam modelos experimentais com roedores, não avaliavam comportamentos relacionados à ansiedade ou ao medo, ou não abordavam diretamente os processos de extinção ou reconsolidação de memórias aversivas.
Resultados e Discussão
Efeitos do CBD sobre a extinção de memórias aversivas
A TABELA 1 contém os detalhes metodológicos dos estudos que avaliaram esta questão em animais de laboratório. Os resultados sugerem que o CBD (10 mg/kg) pode facilitar a aquisição da extinção do medo após administração sistêmica e aguda em ratos, isto é, há redução adicional do tempo de congelamento relativo ao controle. A faixa de dose na qual o CBD facilitaria a extinção do medo após administração sistêmica e repetida ainda não foi estabelecida. Entretanto, já foi relatado em um protocolo com três sessões de aquisição da extinção que a infusão de CBD nos ventrículos cerebrais antes de cada sessão foi capaz de potencializar a extinção da resposta de medo condicionado[26]. Esse efeito foi prevenido pelo pré-tratamento com um antagonista dos receptores CB1, mas não com um antagonista dos canais do receptor de potencial transitório vanilóide 1 (TRPV1), sugerindo que o CBD aceleraria a extinção de memórias aversivas via ativação de receptores CB1[26].
Os receptores CB1 têm expressão relativamente alta em terminais GABAérgicos pré-sinápticos[27,28] do córtex infralímbico[29], uma área fundamental no processo de extinção de memórias aversivas[30-32]. Esse receptor está acoplado a uma proteína G inibitória e, quando ativado, aumenta as correntes de K+ e inibe a atividade da adenilil ciclase, reduzindo os níveis de AMPc e inibindo os canais de cálcio[27,28,33]. Assim, a redução da inibição GABAérgica via ativação dos receptores CB1 ocasiona um aumento da atividade neural no córtex infralímbico, o que embasaria a facilitação da extinção do medo após a infusão local de CBD, e explicaria a ausência desse efeito pela administração sistêmica prévia de um antagonista dos receptores CB1[34].
Na verdade, a administração sistêmica aguda de CBD pode afetar a aquisição da extinção de memórias aversivas contextuais de maneira bidirecional, dependendo da intensidade do condicionamento empregado: quando o condicionamento é fraco, o CBD prejudica a extinção; quando ele é forte, o CBD facilita a extinção[35]. Esses resultados sugerem que o efeito do CBD na extinção depende da intensidade da experiência aversiva dos animais durante o condicionamento, fato que nunca havia sido relatado antes e cujos mecanismos subjacentes não estão claros, mas parecem envolver a hiperativação de glicocorticoides[35].
Diferente do observado com memórias aversivas contextuais, quando se utilizou o condicionamento aversivo olfatório com odor de predador para induzir memórias aversivas, o tratamento sistêmico com CBD não alterou a expressão do comportamento de congelamento, ou seja, não afetou o processo de extinção[36] Neste trabalho, a retenção da memória de extinção não foi avaliada, dificultando a interpretação completa do resultado, uma vez que o CBD poderia ter alterado esse parâmetro mesmo sem interferir com a aquisição. A dose de CBD administrada nesse estudo (5,0 mg/kg) está entre as mais baixas da curva em “U” invertido para os efeitos do CBD, que varia de 3,0 a 30 mg/kg no caso da reconsolidação[37]. O uso de uma dose relativamente baixa, um intervalo curto entre a injeção sistêmica e o teste, e o fato de se tratar de uma memória remota, que tende a ser mais resistente à extinção, poderiam explicar a ausência de efeitos do CBD[36].
A administração sistêmica de CBD não alterou a extinção em animais submetidos ao condicionamento aversivo auditório[38], um desfecho que difere daquele obtido por Song et al.[35], que utilizaram protocolo, animais, dose e via de administração semelhantes. Por outro lado, os grupos tratados com CBD expressaram menores níveis de congelamento no teste de recuperação espontânea, realizado pelo menos 20 dias após a extinção, sugerindo um potencial efeito do CBD na redução do medo a longo prazo[38].
Até 2023, todos os estudos que avaliaram os efeitos do CBD na extinção do medo foram realizados com roedores machos[26,34-36,38], o que gerava uma lacuna na literatura. Considerando a maior suscetibilidade das mulheres a transtornos relacionados à ansiedade e ao estresse em comparação aos homens, a evidência científica limitada sobre como o CBD influencia esse processo no sexo feminino poderia ter implicações terapêuticas[39]. Assim, o trabalho realizado por Franzen et al.[40] focou na compreensão dos efeitos do CBD em ratas. Não houve diferenças estatisticamente significativas entre os grupos ao longo do ciclo estral[40], corroborando estudos prévios[41,42] e descartando a possibilidade de que a variação hormonal das ratas poderia influenciar a expressão de congelamento neste protocolo. Mais importante, o grupo de ratas submetidas ao condicionamento aversivo contextual e tratadas com CBD por via intraperitoneal um dia antes da extinção expressou níveis de congelamento significativamente menores do que as do grupo controle no início da sessão[40]. Esse resultado sugere um prejuízo na reconsolidação após a reativação e desestabilização da memória, levando à redução do congelamento na sessão de extinção, semelhante ao observado nos trabalhos com machos[37,43-46].
No condicionamento aversivo contextual, as respostas desencadeadas pela exposição ao EC dependem da atividade e plasticidade do hipocampo dorsal[47]. A infusão de CBD no hipocampo dorsal facilitou o processo de extinção durante a sessão de aquisição, mas os níveis de congelamento no teste para avaliar a retenção da memória de extinção permaneceram inalterados, resultado equivalente ao observado após administração sistêmica. Isso indica que a administração local de CBD é capaz de desencadear tais efeitos mnemônicos, resultando na redução dos níveis de congelamento em ratas[40].
Em resumo, os resultados apresentados na TABELA 1 sugerem que o CBD pode potencializar a extinção de memórias aversivas em ratos e ratas por meio da ativação de receptores CB1. No entanto, os efeitos variam conforme a linhagem, a dose, a via de administração, o intervalo entre a administração e o teste, bem como os protocolos experimentais empregados. Isso sugere que o CBD pode ser uma ferramenta útil na modulação de memórias aversivas, mas que a eficácia do tratamento dependeria do contexto experimental. Até o momento, os estudos publicados (TABELA 1) não fornecem evidências suficientes para inferirmos quais áreas encefálicas estão envolvidas e quais são os mecanismos neurais subjacentes aos efeitos do CBD sobre a extinção de memórias aversivas. Além disso, possíveis diferenças entre espécies e sexo precisam ser avaliadas sistematicamente, visto que os estudos utilizaram majoritariamente ratos machos, sendo ainda escassos os resultados em camundongos e fêmeas. Além disso, do ponto de vista translacional, seria pertinente avaliar os efeitos do CBD em protocolos que mimetizam memórias traumáticas, ou seja, as que apresentam déficits de extinção e são mais generalizadas[48]. Também seria adequado complementar os resultados existentes com os obtidos em protocolos de esquiva, o que possibilitaria obtermos um panorama mais amplo acerca dos efeitos do CBD no processo de extinção de memórias aversivas.
Efeitos do CBD sobre a reconsolidação de memórias aversivas
A TABELA 2 contém os detalhes metodológicos dos estudos que avaliaram esta questão em animais de laboratório. Os resultados sugerem que o CBD (3,0-50 mg/kg) pode prejudicar a reconsolidação de memórias aversivas, isto é, sua administração aguda e sistêmica imediatamente após uma breve exposição ao EC (para evocar e desestabilizar a memória) reduz persistentemente (≥ uma semana) o tempo de congelamento em ratos, ratas e camundongos[37,43-46,49,50]. Por outro lado, o CBD não alterou a reconsolidação quando foi administrado seis horas após a sessão de evocação e desestabilização, indicando que esse efeito seria causado apenas dentro da janela de reestabilização da memória[37,44,45]. De forma semelhante, não houve influência do ciclo estral sobre os prejuízos induzidos pelo CBD na reconsolidação de memórias aversivas[50].
Quanto à neurobiologia desses efeitos do CBD, o estudo de Bayer et al.[44] relatou que a infusão de CBD no córtex cingulado anterior ou pré-límbico, mas não no infralímbico, foi suficiente para prejudicar a reconsolidação de memórias aversivas reativadas em ratos. Esses achados se alinham com os resultados de análises imuno-histoquímicas da proteína Zif268/Egr-1, um marcador do processo de reconsolidação, embora eles não sejam totalmente compatíveis com a observação de que a ativação dos receptores CB1 localizados nessas três sub-regiões do córtex pré-frontal medial seria necessária[44]. Assim, a atividade e a plasticidade nos córtices cingulado anterior e pré-límbico seriam essenciais para a reconsolidação de memórias aversivas[51,52], já o córtex infralímbico estaria mais envolvido com a extinção do medo[34].
Franzen et al.[45] relataram que a infusão de CBD (30 pmol) na região CA1 do hipocampo dorsal imediatamente após a reativação prejudicou a reconsolidação de memórias aversivas em ratas, um efeito mediado via receptores CB1. Ressalta-se que a ativação desse receptor pelo CBD seria indireta e associada ao aumento dos níveis do endocanabinoide anandamida[53,54]. O CBD promoveria esse efeito através da inibição da enzima hidrolase de amida de ácido graxo (FAAH), que degrada a anandamida. A inibição farmacológica da FAAH aumenta os níveis de anandamida no hipocampo e produz efeitos sobre a reconsolidação de memórias aversivas semelhantes aos do CBD[25,55]. Além disso, o CBD também pode interferir com o transporte intracelular de anandamida, competindo com proteínas de ligação a ácidos graxos, o que contribuiria para o aumento dos níveis desse endocanabinoide[56]. Outro mecanismo possível seria o aumento da síntese de anandamida pela ativação da enzima N-acil fosfatidiletanolamina fosfolipase D[57].
A administração aguda e sistêmica de CBD (10 mg/kg) imediatamente após a evocação de uma memória aversiva potencializada pela ioimbina foi incapaz de prejudicar a reconsolidação em ratos[43]. A ioimbina aumenta o tônus noradrenérgico no sistema nervoso central e, assim, torna a memória mais intensa e generalizada. Em outro protocolo, ratos e ratas foram submetidas a um condicionamento relativamente mais intenso do que o normal, e a administração da dose supracitada de CBD também não produziu alterações significativas em relação ao grupo controle[50]. Em ambos os casos, no entanto, a administração prévia de D-cicloserina, um agonista parcial dos receptores NMDA para glutamato, foi capaz de potencializar a desestabilização da memória e, assim, possibilitou que o CBD prejudicasse a reconsolidação[43,50].
A administração repetida de CBD tem sido testada em modelos de exposição crônica ao estresse, com resultados promissores na atenuação dos sintomas relacionados ao transtorno de estresse pós-traumático, como comportamento de esquiva e respostas emocionais excessivas a estímulos aversivos. Isso sugere que além de seu impacto na reconsolidação, o CBD poderia melhorar a resiliência ao estresse prolongado. Neste contexto, o estudo de Han et al.[46] comparou os efeitos do CBD aos da sertralina, um inibidor seletivo da recaptação de serotonina. Os resultados sugerem que o CBD teria eficácia semelhante à da sertralina, mas possivelmente com menos efeitos colaterais.
Em resumo, o CBD pode prejudicar a reconsolidação de memórias aversivas em ratos, ratas e camundongos, um efeito mediado via ativação do receptor CB1 no hipocampo dorsal, córtex cingulado anterior e/ou córtex pré-límbico. É adequado ressaltar que se combinado com a D-cicloserina, essa propriedade do CBD é mantida mesmo em protocolos que mimetizam memórias traumáticas, ou seja, as que apresentam uma desestabilização insuficiente[48]. Por outro lado, o mecanismo e o local de ação do CBD nesses casos ainda estão em análise. Por fim, seria adequado complementar os resultados existentes utilizando protocolos de esquiva para obtenção de evidências adicionais acerca dos efeitos do CBD sobre a reconsolidação de memórias aversivas.
Efeitos do CBD sobre a expressão de medo e ansiedade
A TABELA 3 contém os detalhes metodológicos dos estudos que avaliaram esta questão em animais de laboratório. Os resultados obtidos em diferentes testes e modelos experimentais são consistentes, mostrando que ratos e camundongos tratados com CBD manifestam menos comportamentos defensivos em resposta a estímulos inatos ameaçadores ou àqueles associados com experiências emocionais desagradáveis. De fato, após administração sistêmica única ou repetida, o efeito ansiolítico do CBD é observado sobretudo em doses intermediárias (1,0-30 mg/kg), mas não em doses mais baixas[59,60] ou mais altas[59,61-66], embora existam alguns resultados conflitantes na literatura quando comportamentos relacionados à ansiedade foram avaliados em camundongos em abstinência espontânea de etanol [67] ou modificados geneticamente [68-70].
O padrão de curva dose-resposta em forma de sino (ou “U” invertido) do CBD pode estar associado à modulação de diferentes alvos farmacológicos, pois há evidências de que a ativação de diferentes receptores é necessária para o efeito ansiolítico do CBD. Já foi demonstrado que o antagonismo de receptores 5-HT1A[17,40,61,64,65,70-78], CB1[26,34,79-81] e CB2[82] impede o efeito ansiolítico desse fitocanabinoide. Embora o trabalho de Onaivi et al.[59] sugira que o efeito ansiolítico do CBD em camundongos também depende da ativação de receptores benzodiazepínicos, posteriormente Moreira et al.[83] mostraram que esse padrão de resposta não é observado em ratos, logo o envolvimento desses receptores no efeito ansiolítico do CBD precisa ser reavaliado. Já a ativação do canal TRPV1 parece contribuir para a ausência de efeito do CBD após administração única e em doses mais elevadas[62]. Por outro lado, a dessensibilização do canal TRPV1 após administração crônica de CBD[84] poderia justificar o efeito ansiolítico dessa substância quando doses elevadas foram administradas diariamente por algumas semanas[68,69].
Além da ativação de receptores diferentes, evidências apontam que a administração repetida de CBD modifica em várias regiões encefálicas a expressão de proteínas que contribuem para a regulação de processos neurobiológicos relacionados a alterações emocionais, incluindo o fator neurotrófico derivado do encéfalo (BDNF), fosfo-ERK/2, transportador de serotonina Slc6a4, receptores CB1, CB2, TrkB – cujo agonista é o BDNF – e os para glicocorticoides[67,85,86], bem como facilita a neurogênese hipocampal mediada pela ativação dos receptores CB1 e CB2[82,87]. O efeito ansiolítico após administração única de CBD, por sua vez, já foi associado à redução dos níveis dos neurotransmissores dopamina e serotonina no núcleo accumbens e no córtex pré-frontal medial[88], além da alteração na expressão dos receptores CB1, CB2, Gpr55 e GABAA em regiões do hipocampo, amígdala e núcleo accumbens[67,80], aumento na expressão de BDNF no núcleo accumbens, córtex pré-frontal medial e amígdala basolateral[89], diminuição da expressão das proteínas S100β e Iba1 na amígdala[90] e regulação da expressão de diversos outros genes[91].
Devido à interação do CBD com diferentes alvos celulares, que resultam em alterações moleculares distintas em determinadas regiões encefálicas, estudos relatam que o CBD pode promover efeitos opostos quando injetado diretamente em regiões do circuito encefálico envolvido na regulação da expressão de comportamentos defensivos relacionadas à ansiedade e ao pânico, variando também de acordo com o teste ou paradigma avaliado, bem como com a exposição prévia a estímulos estressores. A infusão de CBD no córtex pré-límbico atenuou a evocação/expressão de memórias formadas no condicionamento aversivo contextual[63,64], promoveu efeito ansiolítico no labirinto em T elevado[77] e no labirinto em cruz elevado em animais submetidos ao estresse de contenção[64], mas promoveu efeito ansiogênico em animais sem estresse prévio[64]. Os efeitos do CBD foram associados à ativação local do receptor 5-HT1A[64,67]. Em outra sub-região do córtex pré-frontal medial, a do infralímbico, uma infusão de CBD facilitou a evocação/expressão de memórias aversivas[63,65], e promoveu efeito ansiolítico no labirinto em cruz elevado apenas em animais não estressados[65]. Já a administração repetida de CBD no córtex infralímbico atenuou a evocação/expressão de memórias aversivas[34]. A ativação local do receptor 5-HT1A também estaria envolvida nesses efeitos do CBD[65].
A infusão de CBD na substância cinzenta periaquedutal dorsolateral reduz os comportamentos de fuga (efeito panicolítico) durante a estimulação elétrica dessa região encefálica[17], além de promover efeito ansiolítico em outros testes de ansiedade[17,61,62], efeitos relacionados à ativação dos receptores 5-HT1A[17,61,75]. No núcleo do leito da estria terminal, a ativação dos receptores 5-HT1A contribui para o efeito ansiolítico promovido pela infusão local de CBD[71,74] observado em animais testados do labirinto em cruz elevado e no teste de Vogel[72], enquanto na amígdala central tal efeito foi observado apenas no teste do campo aberto[92]. A infusão de CBD no núcleo do leito da estria terminal também afetou a evocação/expressão de memórias aversivas[74], já a infusão intra-núcleo ventral do hipotálamo produziu efeito panicolítico via ativação do receptor CB1, reduzindo o comportamento de fuga após estimulação com NMDA[79]. No hipocampo dorsal, a ativação do receptor 5-HT1A foi associada com a redução do tempo de congelamento após administração sistêmica de CBD[40].
Esses resultados corroboram outros estudos mostrando que regiões do córtex pré-frontal, hipocampo, diencéfalo e mesencéfalo estão envolvidas no circuito neural das respostas de roedores relacionadas ao estresse e à ansiedade frente a estímulos aversivos[93]. Além disso, estão de acordo com evidências que apontam o envolvimento de receptores 5-HT1A e CB1 na manifestação de comportamentos associados com a ansiedade em roedores[94-98]. Além disso, as diferenças observadas após infusão do CBD em determinadas regiões encefálicas e/ou em diferentes testes comportamentais refletem os desafios no estudo dos comportamentos relacionados à ansiedade em roedores, visto que circuitos cerebrais distintos podem ser recrutados para expressão de respostas defensivas inatas do animal (ex.: na presença de um predador ou de um estímulo naturalmente aversivo) ou para respostas condicionadas a eventos estressantes e possivelmente dolorosos, tal como a aplicação de choques elétricos nas patas[99-103]. Apesar dessa ressalva, a maioria dos estudos em roedores (TABELA 3) relata efeitos ansiolíticos do CBD após administração sistêmica ou i.c.v.[26].
Além da neurobiologia envolvida na regulação da expressão de comportamentos defensivos, outra questão a ser considerada é que poucos estudos utilizaram roedores fêmeas e/ou investigaram uma possível influência do sexo sobre o efeito ansiolítico do CBD. Embora os estudos que utilizaram ratas sugiram que o CBD produz efeito ansiolítico em diversos testes e em doses e protocolos experimentais semelhantes aos utilizados em machos[40,88,89,104], já foi relatado que além da exposição prévia a estímulos estressores[60], a fase do ciclo estral pode interferir na resposta comportamental observada, sendo o efeito do CBD observado apenas durante o diestro tardio[104,105]. Além disso, os resultados seriam menos consistentes entre as espécies, pois em camundongos alguns estudos têm relatado ausência de efeitos do CBD na ansiedade[106,107] ou mesmo efeito ansiogênico em fêmeas[90,108]. Assim, a realização de mais estudos pré-clínicos com fêmeas é necessária para esclarecer o efeito do CBD na regulação de processos neurobiológicos relacionados à emocionalidade, sobretudo se forem consideradas as possíveis aplicações terapêuticas do CBD na clínica e a maior suscetibilidade das mulheres a transtornos relacionados à ansiedade[39].
Em resumo, a administração aguda e repetida de CBD promove efeitos ansiolíticos em roedores, sendo influenciados por variáveis experimentais como dose e via de administração, espécie e sexo dos animais, assim como o teste comportamental empregado. Os mecanismos de ação pelos quais o CBD regula as respostas defensivas e os comportamentos relacionados à ansiedade em roedores são complexos e, embora venham sendo extensivamente estudados, ainda não estão completamente elucidados[54]. Até o momento, os estudos mostram que a ativação de receptores 5-HT1A, CB1 e/ou CB2 contribui para a obtenção do efeito ansiolítico do CBD em doses intermediárias, enquanto a ativação do canal TRPV1 parece estar associada à ausência de efeito do CBD em doses altas. Quanto aos circuitos neurais relacionados ao efeito ansiolítico do CBD em roedores, subregiões do córtex pré-frontal medial, hipocampo, amigdala, substância cinzenta periaquedutal, núcleo do leito da estria terminal e núcleo ventral do hipotálamo estariam envolvidas. Entretanto, é importante destacar que testes ou paradigmas que requerem a execução de estratégias defensivas reativas ou de esquiva recrutam regiões e circuitos encefálicos distintos, o que pode justificar diferenças quanto às respostas comportamentais observadas após administração local ou sistêmica de CBD. Dessa forma, um avanço na compreensão da neurobiologia de tais efeitos em modelos animais pode contribuir para a avaliação do potencial terapêutico do CBD no manejo de determinadas condições clínicas em humanos.
Considerações sobre o padrão de resultados e as discrepâncias observadas
A curva dose-resposta em formato de sino ("U" invertido) é uma característica amplamente documentada em estudos pré-clínicos com CBD, incluindo os que focam em ansiedade, extinção e reconsolidação de memórias aversivas. Ou seja, o CBD possui efeitos mais pronunciados em doses intermediárias, enquanto doses muito baixas ou altas podem ser ineficazes. De fato, Guimarães et al.[109] testaram ratos no labirinto em cruz elevado e mostraram que a administração sistêmica aguda de CBD produz uma curva em formato de sino, sendo ansiolítico em doses baixas e intermediárias (2,5-10 mg/kg), mas não em doses relativamente altas (20 mg/kg). Embora alguns resultados contraditórios existam na literatura, a maioria dos estudos que utilizam testes de ansiedade ou que investigam os efeitos do CBD em protocolos de condicionamento Pavloviano apresenta o mesmo perfil de efeitos[3]. Campos et al.[73] também relataram que uma dose intermediária de CBD (10 mg/kg) produz um efeito ansiolítico, enquanto doses mais altas (20 mg/kg) ou baixas (1,0 mg/kg) não causaram o mesmo efeito. A ativação de receptores 5-HT1A induzida pelo CBD (em doses intermediárias) seria fundamental para seu efeito ansiolítico, enquanto doses mais altas também ativariam o canal TRPV1, o que atenua essa resposta ansiolítica[61]. Estudos sobre a reconsolidação de memórias aversivas também observaram que uma dose intermediária de CBD (10 mg/kg) seria mais eficaz do que as de 3,0 e 30 mg/kg[37]. O perfil em "U" invertido nesses estudos indica que doses intermediárias de CBD produziram a interferência máxima sobre a reconsolidação de memórias aversivas.
Os mecanismos subjacentes a essa curva dose-resposta em "U" invertido podem estar relacionados à ativação e modulação de alvos farmacológicos diferentes. Evidências sugerem que a ativação de receptores específicos é necessária para que o CBD exerça seu efeito ansiolítico. Por exemplo, o antagonismo de receptores 5-HT1A impede esse efeito[17,40,61,64,65,70-74,76-78]. O CBD também interage com outros alvos farmacológicos, tais como os receptores TRPV1, CB1 e CB2. Em doses baixas, pode não haver ativação suficiente desses receptores para induzir efeitos significativos. Por outro lado, doses maiores de CBD podem ativar o canal TRPV1, que exerce uma modulação negativa sobre a ansiedade[75]. Assim, a seleção de uma dose adequada é essencial para obtenção dos efeitos desejáveis do CBD. Esse padrão de resposta em "U" invertido tem implicações importantes para estudos pré-clínicos e futuros ensaios clínicos, destacando a necessidade de definir com precisão as faixas de doses apropriadas para cada aplicação terapêutica.
Uma rede complexa de áreas encefálicas modula a manifestação de respostas defensivas inatas ou aprendidas em roedores. Inúmeros estudos mostram a participação de sub-regiões da substância cinzenta periaquedutal, núcleo do leito da estria terminal, hipocampo, amígdala e córtex pré-frontal medial, conforme a natureza do estímulo apresentado, e que a ativação de um determinado receptor em cada uma delas pode contribuir de maneira diferente para a manifestação de comportamentos relacionados à ansiedade ou ao medo aprendido[100,124]. Isso explicaria, por exemplo, as diferenças comportamentais observadas após a infusão de CBD nos córtices pré-límbico e infralímbico[44,63], pois eles se conectam com circuitos diferentes na amígdala e promovem efeitos opostos sobre a regulação do medo e da ansiedade. O córtex pré-límbico envia projeções excitatórias principalmente para a região basolateral da amígdala, que estimula projeções de saída do núcleo medial da amígdala central para a substância cinzenta ventrolateral, que promove respostas de medo e ansiedade. Já o córtex infralímbico envia projeções excitatórias para as células intercalares (neurônios inibitórios) da amígdala central, que inibem as projeções de saída. Desta forma, a ativação do córtex pré-límbico favorece respostas de medo condicionado e ansiedade, já a ativação do córtex infralímbico atenua sua expressão e/ou facilita a extinção[102,125]. O efeito ansiolítico e ansiogênico do CBD quando administrado diretamente nos córtices pré-límbico e infralímbico, respectivamente, pode estar relacionado à redução da atividade dessas regiões através de sua interação com o receptor 5-HT1A[64,65], que está localizado principalmente em neurônios piramidais no córtex pré-frontal e, quando ativado, reduz as correntes elétricas excitatórias[126-128]. Paradoxalmente, a administração sistêmica de agonistas do receptor 5-HT1A aumenta a atividade dos neurônios piramidais no córtex pré-frontal através da interação preferencial com receptores localizados em interneurônios GABAérgicos, diminuindo sua atividade e efeito inibitório sobre os neurônios piramidais[129]. Logo, a resultante da regulação da atividade celular em populações neuronais distintas nas sub-regiões do córtex pré-frontal medial via ativação do receptor 5-HT1A poderia justificar o efeito ansiolítico observado após administração sistêmica de CBD[130]. Além disso, após administração sistêmica, o CBD ativa o receptor 5-HT1A em outras áreas encefálicas, tais como a substância cinzenta periaquedutal dorsolateral, núcleo do leito da estria terminal, núcleo ventromedial do hipotálamo e hipocampo dorsal, o que contribui para o seu efeito ansiolítico[40,61,72,79].
Outras evidências pré-clínicas apontam que dentre as alterações endócrinas e autonômicas induzidas pela exposição a eventos estressores está o aumento nos níveis endógenos de glicocorticoides e de sua interação com receptores glicocorticoides, o que reduziria a expressão do receptor 5-HT1A em regiões encefálicas envolvidas na regulação das respostas comportamentais relacionadas ao medo e à ansiedade, tais como o córtex pré-frontal e o hipocampo[126,131,132]. Além de corroborar estudos que apontam a participação do receptor 5-HT1A na resiliência ao estresse[133-136], esse mecanismo embasaria a ausência de efeito ansiolítico quando uma única dose de CBD foi administrada em animais submetidos a protocolos de exposição ao predador[53,73] ou estresse crônico imprevisível[60], bem como após administração repetida em animais submetidos a um protocolo de isolamento social prolongado[89]. O efeito ansiogênico da administração de CBD no córtex infralímbico, mediado pela sua interação com o receptor 5-HT1A, também não foi observado em animais previamente submetidos ao estresse de contenção[65].
A administração repetida de CBD em roedores parece atenuar as alterações induzidas pelo estresse e contribuir para a redução dos comportamentos associados à ansiedade e ao medo aprendido[53,73,82,87,120]. Esse efeito estaria associado com uma diminuição da expressão do gene dos receptores glicocorticoides [120], a ativação do receptor 5-HT1A[53,73,136], a ativação e/ou aumento da expressão de receptores CB1 e CB2 [120], assim como ao aumento da neurogênese e plasticidade sináptica no hipocampo induzida pelos receptores canabinoides[82,87,120,137]. Além disso, estudos relatam que a administração crônica de CBD pode restaurar a função serotoninérgica comprometida pelo estresse, visto que ele aumenta a disponibilidade de serotonina em áreas encefálicas como o córtex pré-frontal medial e o hipocampo, facilitando a ativação dos receptores 5-HT1A, cuja expressão pode estar diminuída[138-140]. Assim, além das diferenças observadas após administração do CBD diretamente em áreas encefálicas que contribuem de maneira distinta para a expressão de respostas defensivas, a exposição prévia dos roedores a eventos estressores ocasiona alterações neurais que podem influenciar a ação do CBD, determinando seu efeito ansiolítico, ansiogênico ou a ausência de efeito dependendo do protocolo avaliado.
Vários testes e protocolos experimentais podem ser utilizados para se analisar os efeitos do CBD sobre os comportamentos relacionados ao medo e à ansiedade em animais de laboratório. No caso do medo, protocolos de condicionamento são amplamente empregados para induzir a formação de memórias aversivas e avaliar as respostas defensivas expressas pelos roedores. Embora os resultados sejam robustos e apontem que a administração de CBD atenua as memórias aversivas ao potencializar sua extinção (TABELA 1) ou prejudicar sua reconsolidação (TABELA 2), os resultados podem ser influenciados por vários aspectos, incluindo a intensidade do condicionamento[35] e a dose de CBD testada[36,38]. Neste contexto, os resultados aparentemente conflitantes do CBD sobre a reconsolidação de memórias aversivas podem ser melhor compreendidos considerando-se as características da memória aversiva avaliada (“normal” ou generalizada) e o grau de sua desestabilização após evocação[37,43,45,50]. Ainda, a variabilidade dos efeitos do CBD em paradigmas de memória aversiva também pode estar associada ao tipo de comportamento que é mensurado (congelamento, esquiva ou darting) e à natureza do estímulo condicionado empregado (contexto, som, odor ou luz), devido ao recrutamento de circuitos neurais relativamente diferentes[103,141,142]. Ainda, é importante considerar que machos e fêmeas aprendem e expressam comportamentos defensivos de forma distinta[42,45,141], logo diferenças sexuais também podem influenciar as respostas comportamentais e os efeitos farmacológicos do CBD.
A premissa de que cada teste comportamental é único também valeria para os que avaliam ansiedade e medo[8]. As questões supracitadas atestam as nuances e complexidades de comparação dos resultados de estudos com metodologias e propósitos diferentes. Uma estratégia prudente para elucidar os efeitos do CBD (e de outras substâncias com potencial terapêutico promissor) sobre as memórias aversivas, o medo e a ansiedade é usar um desenho experimental que inclua machos e fêmeas e envolva a análise de comportamentos e respostas autonômicas e endócrinas complementares, além de estudos para descartar possíveis efeitos não específicos sobre a atividade locomotora[143].
Potencial relevância translacional dos resultados em roedores para o tratamento dos transtornos de ansiedade e estresse em humanos
Conforme revisado por Simei et al.[5], pesquisas básicas e clínicas realizadas no Brasil nos últimos 50 anos indicam um efeito ansiolítico do CBD, despertando interesse pelo seu potencial terapêutico como uma opção no tratamento de transtornos de ansiedade. Han et al.[144] realizaram uma revisão sistemática de oito artigos envolvendo um total de 316 participantes, sendo observado um impacto positivo e significativo (com tamanho do efeito grande) do CBD no tratamento dos transtornos de ansiedade generalizada (TAG), ansiedade social (TAS) e estresse pós-traumático (TEPT). Embora esses resultados devam ser interpretados com cautela devido ao tamanho relativamente limitado da amostra clínica, eles vão ao encontro do que tem sido observado em estudos conduzidos com humanos saudáveis[145] e animais de laboratório (TABELA 3). Além disso, os efeitos mnemônicos do CBD (TABELAS 1 e 2) poderiam complementar a sua ação ansiolítica, promovendo benefícios adicionais e mais duradouros[146,147].
Essa revisão focou em estudos com animais de laboratório, que auxiliam no melhor entendimento de aspectos relacionados à extinção e reconsolidação de memórias aversivas ou traumáticas, bem como ao manejo de transtornos relacionados à ansiedade e ao estresse em humanos. Através de estudos pré-clínicos é possível utilizar diferentes protocolos experimentais para identificar intervenções comportamentais e/ou farmacológicas que podem ser testadas em humanos para otimizar os resultados terapêuticos[148,149]. Os protocolos de extinção do medo em roedores, por exemplo, estão associados à terapia de exposição, que é uma abordagem clínica utilizada para reduzir as respostas de medo e ansiedade na qual indivíduos são expostos em um ambiente seguro a pistas relacionadas a um estímulo aversivo ou trauma. As intervenções de reconsolidação em roedores, por sua vez, correspondem às estratégias clínicas exploradas em terapias como as intervenções de atualização da reconsolidação[150]. Em ambos os casos, essas sessões terapêuticas seriam realizadas sobre os efeitos do CBD ou sucedidas pela administração imediata dessa substância. O monitoramento de possíveis reações adversas (ex.: sedação) e a análise dos desfechos de interesse poderiam contar com a ajuda de técnicas de neuroimagem para avaliar mudanças na atividade cerebral relacionadas aos processos subjacentes. Ainda que os desfechos mais avaliados em animais de laboratório (ex.: comportamentos de congelamento, esquiva e avaliação de risco) sejam relativamente diferentes dos que são analisados em humanos, que incluem a resposta de condutância da pele e as alterações no reflexo de sobressalto medidas através de eletromiografia, e medidas subjetivas dos níveis de medo, ansiedade ou desconforto utilizando determinadas escalas validadas, há translação suficiente dos resultados em roedores para os humanos, o que encoraja trabalhos futuros.
Conclusões e sugestões para estudos futuros
Um número crescente de estudos científicos em animais de laboratório indica que o CBD pode influenciar o “destino” das memórias aversivas, já que ele prejudicaria a reconsolidação após a evocação delas. O CBD também pode acelerar a extinção de memórias aversivas e atenuar a expressão de comportamentos defensivos relacionados ao medo e à ansiedade, embora existam resultados conflitantes. Essas propriedades farmacológicas são relevantes para o tratamento de transtornos de ansiedade e estresse em humanos[2,151]. No entanto, seria adequado validar essas evidências em protocolos que induzem memórias com características traumáticas e respostas de medo/ansiedade inapropriadas em ratos e camundongos[43,48,50]. Além disso, três outras lacunas de conhecimento para um entendimento mais aprofundado dos efeitos do CBD são as diferenças sexuais, a suscetibilidade à interferência conforme as memórias aversivas/traumáticas envelhecem e se generalizam, e os efeitos de regimes de tratamento repetidos.
Os efeitos agudos do CBD decorrem de alterações na função dos sistemas de neurotransmissão da serotonina (via receptor 5-HT1A) e da anandamida (via receptores CB1 e CB2). No entanto, seus efeitos após tratamento repetido podem envolver mecanismos ou alterações adicionais e mais graduais, tais como aumento da neuroplasticidade e modificações epigenéticas[54,152,153]. Uma lacuna significativa na nossa compreensão dos efeitos psicotrópicos do CBD reside em estabelecer uma conexão causal entre suas ações neurobiológicas e seus efeitos comportamentais. Com vários alvos em potencial (FIGURA 4), nem sempre se compreende quais mecanismos embasam os desfechos observados em cada situação. Portanto, é essencial complementar os estudos comportamentais focados em seus efeitos mnemônicos e emocionais com pesquisas que abordem os mecanismos celulares e moleculares supostamente implicados.
: Alvos farmacológicos associados aos efeitos do canabidiol (CBD) sobre a possível facilitação da extinção do medo, prejuízo da reconsolidação de memórias aversivas e redução da expressão do medo e ansiedade em animais de laboratório, conforme dados compilados nas TABELAS 1, 2 e 3.
Evidências experimentais indicam que o CBD pode modular a atividade glial, especialmente da micróglia e dos astrócitos, que desempenham papéis fundamentais nas respostas neuroimunes, na plasticidade sináptica e na neuroproteção[154-156]. Embora a contribuição desses efeitos sobre a glia para as ações emocionais e mnemônicas do CBD revisadas aqui ainda não tenha sido investigada sistematicamente, estudos que avaliaram os efeitos do CBD em modelos animais de esquizofrenia[157], epilepsia[158], e depressão[159] corroboram a relevância dessa análise. Xie et al.[160] usaram um protocolo modificado de estresse prolongado único em camundongos para induzir um fenótipo semelhante ao TEPT. A administração de CBD antes das avaliações comportamentais inibiu os comportamentos relacionados ao TEPT e reverteu as alterações na morfologia de micróglias/macrófagos no córtex pré-frontal e no hipocampo ao inibir a neuroinflamação por meio de sua ação nos receptores CB2. Com base nos estudos supracitados, é plausível que a modulação da atividade ou função glial contribua para os efeitos do CBD sobre a ansiedade e a memória aversiva..
Agradecimentos
Ao Portal de Periódicos da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pelo acesso gratuito a artigos científicos.
Referências
- 1 Guimarães FS. Historical perspective on the therapeutic potential of cannabidiol. Int Rev Neurobiol. 2024; 177:1-9. [ https://doi.org/10.1016/bs.irn.2024.03.008 ].
» https://doi.org/10.1016/bs.irn.2024.03.008 - 2 Lisboa SF, Stern CAJ, Gazarini L, Bertoglio LJ. Cannabidiol effects on fear processing and implications for PTSD: Evidence from rodent and human studies. Int Rev Neurobiol. 2024; 177: 235-250. [ https://doi.org/10.1016/bs.irn.2024.03.007 ].
» https://doi.org/10.1016/bs.irn.2024.03.007 - 3 Lee JLC, Bertoglio LJ, Guimarães FS, Stevenson CW. Cannabidiol regulation of emotion and emotional memory processing: relevance for treating anxiety-related and substance abuse disorders. Br J Pharmacol. 2017 Oct; 174(19): 3242-3256. [ https://doi.org/10.1111/bph.13724 ].
» https://doi.org/10.1111/bph.13724 - 4 Stern CAJ , de Carvalho CR , Bertoglio LJ , Takahashi RN . Effects of Cannabinoid Drugs on Aversive or Rewarding Drug-Associated Memory Extinction and Reconsolidation. Neuroscience. 2018 Feb 1; 370: 62-80. [ https://doi.org/10.1016/j.neuroscience.2017.07.018 ].
» https://doi.org/10.1016/j.neuroscience.2017.07.018 - 5 Simei JLQ, de Souza JDR, Lisboa JR, Guimarães FS, Crippa JAS. Cannabidiol in anxiety disorders: Current and future perspectives. Int Rev Neurobiol. 2024; 177: 205-234. [ https://doi.org/10.1016/bs.irn.2024.05.003 ].
» https://doi.org/10.1016/bs.irn.2024.05.003 - 6 Campos AC, Fogaça MV, Aguiar DC, Guimarães FS. Animal models of anxiety disorders and stress. Braz J Psychiatry. 2013; 35 Suppl 2: S101-11. [ https://doi.org/10.1590/1516-4446-2013-1139 ].
» https://doi.org/10.1590/1516-4446-2013-1139 - 7 McNaughton N, Corr PJ. A two-dimensional neuropsychology of defense: fear/anxiety and defensive distance. Neurosci Biobehav Rev. 2004 May; 28(3): 285-305. [ https://doi.org/10.1016/j.neubiorev.2004.03.005 ].
» https://doi.org/10.1016/j.neubiorev.2004.03.005 - 8 Carobrez AP , Bertoglio LJ . Ethological and temporal analyses of anxiety-like behavior: the elevated plus-maze model 20 years on. Neurosci Biobehav Rev. 2005; 29(8): 1193-205. [ https://doi.org/10.1016/j.neubiorev.2005.04.017 ].
» https://doi.org/10.1016/j.neubiorev.2005.04.017 - 9 Bourin M, Hascoët M. The mouse light/dark box test. Eur J Pharmacol. 2003 Feb 28; 463(1-3): 55-65. [ https://doi.org/10.1016/s0014-2999 (03)01274-3].
» https://doi.org/10.1016/s0014-2999 (03)01274-3 - 10 File SE, Seth P. A review of 25 years of the social interaction test. Eur J Pharmacol. 2003 Feb 28; 463(1-3): 35-53. [ https://doi.org/10.1016/s0014-2999 (03)01273-1].
» https://doi.org/10.1016/s0014-2999 (03)01273-1 - 11 Millan MJ, Brocco M. The Vogel conflict test: procedural aspects, gamma-aminobutyric acid, glutamate and monoamines. Eur J Pharmacol. 2003 Feb 28; 463(1-3): 67-96. [ https://doi.org/10.1016/s0014-2999 (03)01275-5].
» https://doi.org/10.1016/s0014-2999 (03)01275-5 - 12 Samuels BA, Hen R. Novelty-Suppressed Feeding in the Mouse. In: T. Gould (Eds.), Mood and Anxiety Related Phenotypes in Mice. Neuromethods 2011; 63: 107-21. Humana Press. [ https://doi.org/10.1007/978-1-61779-313-4_7 ].
» https://doi.org/10.1007/978-1-61779-313-4_7 - 13 De Brouwer G, Fick A, Harvey BH, Wolmarans W. A critical inquiry into marble-burying as a preclinical screening paradigm of relevance for anxiety and obsessive-compulsive disorder: Mapping the way forward. Cogn Affect Behav Neurosci. 2019 Feb; 19(1): 1-39. [ https://doi.org/0.3758/s13415-018-00653-4 ].
» https://doi.org/0.3758/s13415-018-00653-4 - 14 Prut L, Belzung C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. Eur J Pharmacol. 2003 Feb 28; 463(1-3): 3-33. [ https://doi.org/10.1016/s0014-2999 (03)01272-x].
» https://doi.org/10.1016/s0014-2999 (03)01272-x - 15 Dielenberg RA, McGregor IS. Defensive behavior in rats towards predatory odors: a review. Neurosci Biobehav Rev. 2001 Dec; 25(7-8): 597-609. [ https://doi.org/10.1016/s0149-7634 (01)00044-6].
» https://doi.org/10.1016/s0149-7634 (01)00044-6 - 16 Junior Zangrossi H , Graeff FG . Serotonin in anxiety and panic: contributions of the elevated T-maze. Neurosci Biobehav Rev. 2014 Oct; 46 (Pt 3): 397-406. [ https://doi.org/10.1016/j.neubiorev.2014.03.007 ].
» https://doi.org/10.1016/j.neubiorev.2014.03.007 - 17 Soares VP , Campos AC , Bortoli VC , Junior Zangrossi H , Guimarães FS , Zuardi AW . Intra-dorsal periaqueductal gray administration of cannabidiol blocks panic-like response by activating 5-HT1A receptors. Behav Brain Res. 2010; 213(2): 225-229. [ https://doi.org/10.1016/j.bbr.2010.05.004 ].
» https://doi.org/10.1016/j.bbr.2010.05.004 - 18 Izquierdo I , Furini CR , Myskiw JC . Fear Memory. Physiol Rev. 2016 Apr; 96(2): 695-750. [ https://doi.org/10.1152/physrev.00018.2015 ].
» https://doi.org/10.1152/physrev.00018.2015 - 19 Kida S. Interaction between reconsolidation and extinction of fear memory. Brain Res Bull. 2023 Apr; 195: 141-144. [ https://doi.org/10.1016/j.brainresbull.2023.02.009 ].
» https://doi.org/10.1016/j.brainresbull.2023.02.009 - 20 Bouton ME , Maren S , McNally GP . Behavioral and Neurobiological Mechanisms of Pavlovian and Instrumental Extinction Learning. Physiol Rev. 2021 Apr 1; 101(2) :611-681. [ https://doi.org/10.1152/physrev.00016.2020 ].
» https://doi.org/10.1152/physrev.00016.2020 - 21 Lee JLC, Nader K, Schiller D. An Update on Memory Reconsolidation Updating. Trends Cogn Sci. 2017 Jul; 21(7): 531-545. [ https://doi.org/10.1016/j.tics.2017.04.006 ].
» https://doi.org/10.1016/j.tics.2017.04.006 - 22 Walsh KH, Das RK, Saladin ME, Kamboj SK. Modulation of naturalistic maladaptive memories using behavioural and pharmacological reconsolidation-interfering strategies: a systematic review and meta-analysis of clinical and 'sub-clinical' studies. Psychopharmacol. 2018 Sep; 235(9): 2507-2527. [ https://doi.org/10.1007/s00213-018-4983-8 ].
» https://doi.org/10.1007/s00213-018-4983-8 - 23 Bandelow B , Baldwin D , Abelli M , Altamura C , Dell'Osso B , Domschke K , et al . Biological markers for anxiety disorders, OCD and PTSD - a consensus statement. Part I: Neuroimaging and genetics. World J Biol Psychiatry. 2016 Aug; 17(5): 321-65. [ https://doi.org/10.1080/15622975.2016.1181783 ].
» https://doi.org/10.1080/15622975.2016.1181783 - 24 Bandelow B et al. Biological markers for anxiety disorders, OCD and PTSD: A consensus statement. Part II: Neurochemistry, neurophysiology and neurocognition. World J Biol Psychiatry. 2017 Apr; 18(3): 162-214. [ https://doi.org/10.1080/15622975.2016.1190867 ].
» https://doi.org/10.1080/15622975.2016.1190867 - 25 Raymundi AM, da Silva TR, Sohn JMB, Bertoglio LJ, Stern CA. Effects of ? 9 -tetrahydrocannabinol on aversive memories and anxiety: a review from human studies. BMC Psychiatry. 2020 Aug 26; 20(1): 420. [ https://doi.org/10.1186/s12888-020-02813-8 ].
» https://doi.org/10.1186/s12888-020-02813-8 - 26 Bitencourt RM, Pamplona FA, Takahashi RN. Facilitation of contextual fear memory extinction and anti-anxiogenic effects of AM404 and cannabidiol in conditioned rats. Eur Neuropsychopharmacol. 2008 Dec; 18(12): 849-59. [ https://doi.org/10.1016/j.euroneuro.2008.07.001 ].
» https://doi.org/10.1016/j.euroneuro.2008.07.001 - 27 Bodor AL, et al. Endocannabinoid signaling in rat somatosensory cortex: laminar differences and involvement of specific interneuron types. J Neurosci. 2005 Jul 20; 25(29): 6845-56. [ https://doi.org/10.1523/JNEUROSCI.0442-05.2005 ].
» https://doi.org/10.1523/JNEUROSCI.0442-05.2005 - 28 Wedzony K, Chocyk A. Cannabinoid CB1 receptors in rat medial prefrontal cortex are colocalized with calbindin- but not parvalbumin- and calretinin-positive GABA-ergic neurons. Pharmacol Rep. 2009 Nov-Dec; 61(6): 1000-7. [ https://doi.org/10.1016/s1734-1140 (09)70161-6].
» https://doi.org/10.1016/s1734-1140 (09)70161-6 - 29 Tsou K, Brown S, Sañudo-Peña MC, Mackie K, Walker JM. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience. 1998 Mar; 83(2): 393-411. [ https://doi.org/10.1016/s0306-4522 (97)00436-3].
» https://doi.org/10.1016/s0306-4522 (97)00436-3 - 30 Vidal-Gonzalez I, Vidal-Gonzalez B, Rauch SL, Quirk GJ. Microstimulation reveals opposing influences of prelimbic and infralimbic cortex on the expression of conditioned fear. Learn Mem. 2006 Nov-Dec; 13(6): 728-33. [ https://doi.org/10.1101/lm.306106 ].
» https://doi.org/10.1101/lm.306106 - 31 Laurent V, Westbrook RF. Inactivation of the infralimbic but not the prelimbic cortex impairs consolidation and retrieval of fear extinction. Learn Mem. 2009 Aug 25; 16(9): 520-9. [ https://doi.org/10.1101/lm.1474609 ].
» https://doi.org/10.1101/lm.1474609 - 32 Sierra-Mercado D , Padilla-Coreano N , Quirk G . Dissociable Roles of Prelimbic and Infralimbic Cortices, Ventral Hippocampus, and Basolateral Amygdala in the Expression and Extinction of Conditioned Fear. Neuropsychopharmacol. 2011; 36: 529-538. [ https://doi.org/10.1038/npp.2010.184 ].
» https://doi.org/10.1038/npp.2010.184 - 33 Földy C, Neu A, Jones MV, Soltesz I. Presynaptic, activity-dependent modulation of cannabinoid type 1 receptor-mediated inhibition of GABA release. J Neurosci. 2006 Feb 1; 26(5): 1465-9. [ https://doi.org/10.1523/JNEUROSCI.4587-05.2006 ].
» https://doi.org/10.1523/JNEUROSCI.4587-05.2006 - 34 Do Monte FH , Souza RR , Bitencourt RM , Kroon JA , Takahashi RN . Infusion of cannabidiol into infralimbic cortex facilitates fear extinction via CB1 receptors. Behav Brain Res. 2013 Aug 1; 250:23-7. [ https://doi.org/10.1016/j.bbr.2013.04.045 ].
» https://doi.org/10.1016/j.bbr.2013.04.045 - 35 Song C , Stevenson CW , Guimaraes FS , Lee JL . Bidirectional Effects of Cannabidiol on Contextual Fear Memory Extinction. Front Pharmacol. 2016 Dec 16; 7: 493. [ https://doi.org/10.3389/fphar.2016.00493 ].
» https://doi.org/10.3389/fphar.2016.00493 - 36 Shallcross J , Hámor P , Bechard AR , Romano M , Knackstedt L & Schwendt M . The Divergent Effects of CDPPB and Cannabidiol on Fear Extinction and Anxiety in a Predator Scent Stress Model of PTSD in Rats. Front Behav Neurosci. 2019; 13: 91. [ https://doi.org/10.3389/fnbeh.2019.00091 ].
» https://doi.org/10.3389/fnbeh.2019.00091 - 37 Stern CA , Gazarini L , Takahashi RN , Guimarães FS , Bertoglio LJ . On disruption of fear memory by reconsolidation blockade: evidence from cannabidiol treatment. Neuropsychopharmacol. 2012; 37(9): 2132-2142. [ https://doi.org/10.1038/npp.2012.63 ].
» https://doi.org/10.1038/npp.2012.63 - 38 Papagianni EP , Warren WG , Cassaday HJ , Stevenson CW . Cannabidiol Prevents Spontaneous Fear Recovery after Extinction and Ameliorates Stress-Induced Extinction Resistance. Int J Mol Sci. 2022 Aug 19; 23(16): 9333. [ https://doi.org/10.3390/ijms23169333 ].
» https://doi.org/10.3390/ijms23169333 - 39 Penninx BW, Pine DS, Holmes EA & Reif A. Anxiety disorders. Lancet. 2021; 397(10277): 914–927. [https://doi.org/10.1016/S0140-6736(21)00359-7].
» https://doi.org/10.1016/S0140-6736(21)00359-7 - 40 Franzen JM, et al. Cannabidiol attenuates fear memory expression in female rats via hippocampal 5-HT1A but not CB1 or CB2 receptors. Neuropharmacol. 2022; 223: 109316. [ https://doi.org/10.1016/j.neuropharm.2022.109316 ]
» https://doi.org/10.1016/j.neuropharm.2022.109316 - 41 da Silva TR, Takahashi RN, Bertoglio LJ, Andreatini R, Stern CA. Evidence for an expanded time-window to mitigate a reactivated fear memory by tamoxifen. Eur Neuropsychopharmacol. 2016 Oct; 26(10): 1601-9. [ https://doi.org/10.1016/j.euroneuro.2016.08.005 ].
» https://doi.org/10.1016/j.euroneuro.2016.08.005 - 42 Franzen JM, Giachero M, Bertoglio LJ. Dissociating retrieval-dependent contextual aversive memory processes in female rats: Are there cycle-dependent differences? Neuroscience. 2019; 406: 542-553. [ https://doi.org/10.1016/j.neuroscience.2019.03.035 ].
» https://doi.org/10.1016/j.neuroscience.2019.03.035 - 43 Gazarini L, Stern CA, Piornedo RR, Takahashi RN, Bertoglio LJ. PTSD-like memory generated through enhanced noradrenergic activity is mitigated by a dual step pharmacological intervention targeting its reconsolidation. Int J Neuropsychopharmacol. 2014; 18(1): pyu026. [ https://doi.org/10.1093/ijnp/pyu026 ].
» https://doi.org/10.1093/ijnp/pyu026 - 44 Bayer H , Stern CAJ , Troyner F , Gazarini L , Guimarães FS , Bertoglio LJ . Medial prefrontal cortex mechanisms of cannabidiol-induced aversive memory reconsolidation impairments. Neuropharmacol. 2022 Mar 1; 205: 108913. [ https://doi.org/10.1016/j.neuropharm.2021.108913 ].
» https://doi.org/10.1016/j.neuropharm.2021.108913 - 45 Franzen JM, Vanz F, Werle I, Guimarães FS, Bertoglio LJ. Cannabidiol impairs fear memory reconsolidation in female rats through dorsal hippocampus CB1 but not CB2 receptor interaction. Eur Neuropsychopharmacol. 2022; 64: 7-18. [ https://doi.org/10.1016/j.euroneuro.2022.08.002 ].
» https://doi.org/10.1016/j.euroneuro.2022.08.002 - 46 Han X, et al. Comparison between cannabidiol and sertraline for the modulation of post-traumatic stress disorder-like behaviors and fear memory in mice. Psychopharmacol. 2022; 239(5): 1605-1620. [ https://doi.org/10.1007/s00213-022-06132-6 ].
» https://doi.org/10.1007/s00213-022-06132-6 - 47 Chen BK, et al. Artificially Enhancing and Suppressing Hippocampus-Mediated Memories. Curr Biol. 2019 Jun 3; 29(11): 1885-1894.e4. [ https://doi.org/10.1016/j.cub.2019.04.065 ].
» https://doi.org/10.1016/j.cub.2019.04.065 - 48 Gazarini L, Stern CAJ, Bertoglio LJ. On making (and turning adaptive to) maladaptive aversive memories in laboratory rodents. Neurosci Biobehav Rev. 2023 Apr; 147: 105101. [ https://doi.org/10.1016/j.neubiorev.2023.105101 ].
» https://doi.org/10.1016/j.neubiorev.2023.105101 - 49 Murkar A , Kent P , Cayer C , James J , Durst T , Merali Z . Cannabidiol and the Remainder of the Plant Extract Modulate the Effects of ?9-Tetrahydrocannabinol on Fear Memory Reconsolidation. Front Behav Neurosci. 2019; 13: 174. [ https://doi.org/10.3389/fnbeh.2019.00174 ].
» https://doi.org/10.3389/fnbeh.2019.00174 - 50 Soares LA, et al. Dual-step pharmacological intervention for traumatic-like memories: implications from D-cycloserine and cannabidiol or clonidine in male and female rats. Psychopharmacol. 2024; 241: 1827-1840. [ https://doi.org/10.1007/s00213-024-06596-8 ].
» https://doi.org/10.1007/s00213-024-06596-8 - 51 Einarsson EO, Nader K. Involvement of the anterior cingulate cortex in formation, consolidation, and reconsolidation of recent and remote contextual fear memory. Learn Mem. 2012; 19 (10): 449-452. [ https://doi.org/10.1101/lm.027227.112 ].
» https://doi.org/10.1101/lm.027227.112 - 52 Stern CA, Gazarini L, Vanvossen AC, Hames MS, Bertoglio LJ. Activity in prelimbic cortex subserves fear memory reconsolidation over time. Learn Mem. 2013; 21(1): 14-20. [ https://doi.org/10.1101/lm.032631.113 ].
» https://doi.org/10.1101/lm.032631.113 - 53 Campos AC , Ferreira FR , Guimarães FS . Cannabidiol blocks long-lasting behavioral consequences of predator threat stress: possible involvement of 5HT1A receptors. J Psychiatric Res. 2012; 46(11): 1501-1510. [ https://doi.org/10.1016/j.jpsychires.2012.08.012 ].
» https://doi.org/10.1016/j.jpsychires.2012.08.012 - 54 Melas PA , Scherma M , Fratta W , Cifani C , Fadda P . Cannabidiol as a Potential Treatment for Anxiety and Mood Disorders: Molecular Targets and Epigenetic Insights from Preclinical Research. Int J Mol Sci. 2021 Feb 13; 22(4): 1863. [ https://doi.org/10.3390/ijms22041863 ].
» https://doi.org/10.3390/ijms22041863 - 55 Stern CAJ, et al. Cannabidiol disrupts the consolidation of specific and generalized fear memories via dorsal hippocampus CB1 and CB2 receptors. Neuropharmacol. 2017; 125: 220-230. [ https://doi.org/10.1016/j.neuropharm.2017.07.024 ].
» https://doi.org/10.1016/j.neuropharm.2017.07.024 - 56 Elmes MW, Kaczocha M, Berger WT, Leung K, Ralph BP, Wang L, et al. Fatty acid-binding proteins (FABPs) are intracellular carriers for ?9-tetrahydrocannabinol (THC) and cannabidiol (CBD). J Biol Chem. 2015; 290(14): 8711-8721. [ https://doi.org/10.1074/jbc.M114.618447 ].
» https://doi.org/10.1074/jbc.M114.618447 - 57 Leishman E , Manchanda M , Thelen R , Miller S , Mackie K , Bradshaw HB . Cannabidiol's Upregulation of N-acyl Ethanolamines in the Central Nervous System Requires N-acyl Phosphatidyl Ethanolamine-Specific Phospholipase D Cannabis. Cannabinoid Res. 2018; 3 (1): 228-241. [ https://doi.org/10.1089/can.2018.0031 ].
» https://doi.org/10.1089/can.2018.0031 - 58 Stern CA, et al. ?9-Tetrahydrocannabinol alone and combined with cannabidiol mitigate fear memory through reconsolidation disruption. Eur Neuropsychopharmacol. 2015; 25(6): 958-965. [ https://doi.org/10.1016/j.euroneuro.2015.02.001 ].
» https://doi.org/10.1016/j.euroneuro.2015.02.001 - 59 Onaivi ES, Green MR, Martin BR. Pharmacological characterization of cannabinoids in the elevated plus maze. J Pharmacol Exp Ther. 1990 Jun; 253(3): 1002-9. [ https://pubmed.ncbi.nlm.nih.gov/2162942/ ].
» https://pubmed.ncbi.nlm.nih.gov/2162942/ - 60 Pérez-Valenzuela E, et al. Sex-Dependent Synergism of an Edible THC: CBD Formulation in Reducing Anxiety and Depressive-like Symptoms Following Chronic Stress. Cur Neuropharmac. 2024; 22(12): 2059-2078. [ https://doi.org/10.2174/1570159X21666230912101441 ].
» https://doi.org/10.2174/1570159X21666230912101441 - 61 Campos AC, Guimarães FS. Involvement of 5HT1A receptors in the anxiolytic-like effects of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacol. 2008; 199(2): 223-230. [ https://doi.org/10.1007/s00213-008-1168-x ].
» https://doi.org/10.1007/s00213-008-1168-x - 62 Campos AC, Guimarães FS. Evidence for a potential role for TRPV1 receptors in the dorsolateral periaqueductal gray in the attenuation of the anxiolytic effects of cannabinoids. Prog Neuropsychopharmacol Biol Psychiatry. 2009; 33(8): 1517-1521. [ https://doi.org/10.1016/j.pnpbp.2009.08.017 ].
» https://doi.org/10.1016/j.pnpbp.2009.08.017 - 63 Lemos JI, Resstel LB, Guimarães FS. Involvement of the prelimbic prefrontal cortex on cannabidiol-induced attenuation of contextual conditioned fear in rats. Behav Brain Res. 2010; 207(1): 105-111. [ https://doi.org/10.1016/j.bbr.2009.09.045 ].
» https://doi.org/10.1016/j.bbr.2009.09.045 - 64 Fogaça MV, Reis FM, Campos AC, Guimarães FS. Effects of intra-prelimbic prefrontal cortex injection of cannabidiol on anxiety-like behavior: involvement of 5HT1A receptors and previous stressful experience. Eur Neuropsychopharmacol. 2014; 24(3): 410-419. [ https://doi.org/10.1016/j.euroneuro.2013.10.012 ].
» https://doi.org/10.1016/j.euroneuro.2013.10.012 - 65 Marinho AL , Vila-Verde C , Fogaça MV , Guimarães FS . Effects of intra-infralimbic prefrontal cortex injections of cannabidiol in the modulation of emotional behaviors in rats: contribution of 5HT1A receptors and stressful experiences. Behav Brain Res. 2015; 286: 49-56. [ https://doi.org/10.1016/j.bbr.2015.02.023 ].
» https://doi.org/10.1016/j.bbr.2015.02.023 - 66 Liu J, Scott BW, Burnham WM. Effects of cannabidiol and ?9-tetrahydrocannabinol in the elevated plus maze in mice. Behav Pharmacol. 2022; 33(2&3): 206-212. [ https://doi.org/10.1097/FBP.0000000000000636 ].
» https://doi.org/10.1097/FBP.0000000000000636 - 67 Gasparyan A , Navarrete F , Navarro D , Manzanares J . Cannabidiol regulates behavioral and brain alterations induced by spontaneous alcohol withdrawal. Neuropharmacol. 2023; 233: 109549. [ https://doi.org/10.1016/j.neuropharm.2023.109549 ].
» https://doi.org/10.1016/j.neuropharm.2023.109549 - 68 Long LE, Chesworth R, Huang XF, McGregor IS, Arnold JC, Karl T. A behavioural comparison of acute and chronic Delta9-tetrahydrocannabinol and cannabidiol in C57BL/6JArc mice. Int J Neuropsychopharmacol. 2010; 13(7): 861-876. [ https://doi.org/10.1017/S1461145709990605 ].
» https://doi.org/10.1017/S1461145709990605 - 69 Long LE, et al. Distinct neurobehavioural effects of cannabidiol in transmembrane domain neuregulin 1 mutant mice. PloS One. 2012; 7(4): e34129. [ https://doi.org/10.1371/journal.pone.0034129 ].
» https://doi.org/10.1371/journal.pone.0034129 - 70 Souza AJ , Guimarães FS , Gomes FV . Cannabidiol attenuates the expression of conditioned place aversion induced by naloxone-precipitated morphine withdrawal through the activation of 5-HT1A receptors. Behav Brain Res. 2023; 450: 114504. [ https://doi.org/10.1016/j.bbr.2023.114504 ].
» https://doi.org/10.1016/j.bbr.2023.114504 - 71 Resstel LB, Tavares RF, Lisboa, SF, Joca SR, Corrêa FM, Guimarães FS. 5-HT1A receptors are involved in the cannabidiol-induced attenuation of behavioural and cardiovascular responses to acute restraint stress in rats. Brit J Pharmacol. 2009; 156(1): 181-188. [ https://doi.org/10.1111/j.1476-5381.2008.00046.x ].
» https://doi.org/10.1111/j.1476-5381.2008.00046.x - 72 Gomes FV, Resstel LB, Guimarães FS. The anxiolytic-like effects of cannabidiol injected into the bed nucleus of the stria terminalis are mediated by 5-HT1A receptors. Psychopharmacol. 2011; 213(2-3): 465-473. [ https://doi.org/10.1007/s00213-010-2036-z ].
» https://doi.org/10.1007/s00213-010-2036-z - 73 Campos AC, Ferreira FR, Guimarães FS. Cannabidiol blocks long-lasting behavioral consequences of predator threat stress: possible involvement of 5HT1A receptors. J Psychiatr Res. 2012 Nov; 46(11): 1501-10. [ https://doi.org/10.1016/j.jpsychires.2012.08.012 ].
» https://doi.org/10.1016/j.jpsychires.2012.08.012 - 74 Gomes FV, Reis DG, Alves FH, Corrêa FM, Guimarães FS, Resstel LB. Cannabidiol injected into the bed nucleus of the stria terminalis reduces the expression of contextual fear conditioning via 5-HT1A receptors. J Psychopharmacol. 2012; 26(1): 104-113. [ https://doi.org/10.1177/0269881110389095 ].
» https://doi.org/10.1177/0269881110389095 - 75 Campos AC, et al. Involvement of serotonin-mediated neurotransmission in the dorsal periaqueductal gray matter on cannabidiol chronic effects in panic-like responses in rats. Psychopharmacol. 2013; 226(1): 13-24. [ https://doi.org/10.1007/s00213-012-2878-7 ].
» https://doi.org/10.1007/s00213-012-2878-7 - 76 Chaves YC, Genaro K, Crippa JA, da Cunha JM, Zanoveli JM. Cannabidiol induces antidepressant and anxiolytic-like effects in experimental type-1 diabetic animals by multiple sites of action. Metab Brain Dis. 2021; 36(4): 639-652. [ https://doi.org/10.1007/s11011-020-00667-3 ].
» https://doi.org/10.1007/s11011-020-00667-3 - 77 Szkudlarek HJ, et al. THC and CBD produce divergent effects on perception and panic behaviours via distinct cortical molecular pathways. Prog Neuropsychopharmacol Biol Psychiatry. 2021; 104: 110029. [ https://doi.org/10.1016/j.pnpbp.2020.110029 ].
» https://doi.org/10.1016/j.pnpbp.2020.110029 - 78 Shu G et al . Cannabidiol exhibits anxiolytic-like effects and antipsychotic-like effects in mice models. Neurosci Lett. 2024; 826: 137723. [ https://doi.org/10.1016/j.neulet.2024.137723 ].
» https://doi.org/10.1016/j.neulet.2024.137723 - 79 Khan AU, et al. Cannabidiol-induced panicolytic-like effects and fear-induced antinociception impairment: the role of the CB1 receptor in the ventromedial hypothalamus. Psychopharmacol. 2020; 237(4): 1063-1079. [ https://doi.org/10.1007/s00213-019-05435-5 ].
» https://doi.org/10.1007/s00213-019-05435-5 - 80 Austrich-Olivares A, García-Gutiérrez MS, Illescas L, Gasparyan A, Manzanares J. Cannabinoid CB1 Receptor Involvement in the Actions of CBD on Anxiety and Coping Behaviors in Mice. Pharmaceuticals. 2022; 15(4): 473. [ https://doi.org/10.3390/ph15040473 ].
» https://doi.org/10.3390/ph15040473 - 81 de Paula Rodrigues BM, Coimbra NC. CB1 receptor signalling mediates cannabidiol-induced panicolytic-like effects and defensive antinociception impairment in mice threatened by Bothrops jararaca lancehead pit vipers. J Psychopharmacol. 2022; 36(12): 1384-1396. [ https://doi.org/10.1177/02698811221115755 ].
» https://doi.org/10.1177/02698811221115755 - 82 Fogaça MV, Campos AC, Coelho LD, Duman RS, Guimarães FS. The anxiolytic effects of cannabidiol in chronically stressed mice are mediated by the endocannabinoid system: Role of neurogenesis and dendritic remodeling. Neuropharmacol. 2018; 135: 22-33. [ https://doi.org/10.1016/j.neuropharm.2018.03.001 ].
» https://doi.org/10.1016/j.neuropharm.2018.03.001 - 83 Moreira FA , Aguiar DC , Guimarães FS . Anxiolytic-like effect of cannabidiol in the rat Vogel conflict test. Prog Neuropsychopharmacol Biol Psychiatry. 2006; 30(8): 1466-1471. [ https://doi.org/10.1016/j.pnpbp.2006.06.004 ].
» https://doi.org/10.1016/j.pnpbp.2006.06.004 - 84 Piao JJ, et al. Cannabidiol Alleviates Chronic Prostatitis and Chronic Pelvic Pain Syndrome via CB2 Receptor Activation and TRPV1 Desensitization. World J Mens Health. 2024; [ https://doi.org/10.5534/wjmh.230352 ].
» https://doi.org/10.5534/wjmh.230352 - 85 ElBatsh MM , Assareh N , Marsden CA , Kendall DA . Anxiogenic-like effects of chronic cannabidiol administration in rats. Psychopharmacol. 2012; 221(2): 239-247. [ https://doi.org/10.1007/s00213-011-2566-z ].
» https://doi.org/10.1007/s00213-011-2566-z - 86 Silva-Cardoso GK, et al. Cannabidiol effectively reverses mechanical and thermal allodynia, hyperalgesia, and anxious behaviors in a neuropathic pain model: Possible role of CB1 and TRPV1 receptors. Neuropharmacol. 2021; 197: 108712. [ https://doi.org/10.1016/j.neuropharm.2021.108712 ].
» https://doi.org/10.1016/j.neuropharm.2021.108712 - 87 Campos AC, et al. The anxiolytic effect of cannabidiol on chronically stressed mice depends on hippocampal neurogenesis: involvement of the endocannabinoid system. Int J Neuropsychopharmacol. 2013; 16(6): 1407-1419. [ https://doi.org/10.1017/S1461145712001502 ].
» https://doi.org/10.1017/S1461145712001502 - 88 Salviato BZ, et al. Female but not male rats show biphasic effects of low doses of ?9-tetrahydrocannabinol on anxiety: can cannabidiol interfere with these effects? Neuropharmacol. 2021; 196: 108684. [ https://doi.org/10.1016/j.neuropharm.2021.108684 ].
» https://doi.org/10.1016/j.neuropharm.2021.108684 - 89 Saad N, Raviv D, Mizrachi Zer-Aviv T, Akirav I. Cannabidiol Modulates Emotional Function and Brain-Derived Neurotrophic Factor Expression in Middle-Aged Female Rats Exposed to Social Isolation. Int J Mol Sci. 2023; 24(20): 15492. [ https://doi.org/10.3390/ijms242015492 ].
» https://doi.org/10.3390/ijms242015492 - 90 Melkumyan M , et al . Effects of cannabidiol, with and without ?9-tetrahydrocannabinol, on anxiety-like behavior following alcohol withdrawal in mice. Front Neurosci. 2024; 18: 1375440. [ https://doi.org/10.3389/fnins.2024.1375440 ].
» https://doi.org/10.3389/fnins.2024.1375440 - 91 Xie G , Qin Y , Wu N , Han X , Li J . Single-Nucleus Transcriptome Profiling from the Hippocampus of a PTSD Mouse Model and CBD-Treated Cohorts. Genes. 2024; 15(4): 519. [ https://doi.org/10.3390/genes15040519 ].
» https://doi.org/10.3390/genes15040519 - 92 Hsiao YT, Yi PL, Li CL, Chang FC. Effect of cannabidiol on sleep disruption induced by the repeated combination tests consisting of open field and elevated plus-maze in rats. Neuropharmacology. 2012; 62(1): 373-384. [ https://doi.org/10.1016/j.neuropharm.2011.08.013 ].
» https://doi.org/10.1016/j.neuropharm.2011.08.013 - 93 Shin LM, Liberzon I. The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacol. 2010; 35(1): 169-191. [ https://doi.org/10.1038/npp.2009.83 ].
» https://doi.org/10.1038/npp.2009.83 - 94 Parks CL , Robinson PS , Sibille E , Shenk T , Toth M . Increased anxiety of mice lacking the serotonin1A receptor. Proc Natl Acad Sci U S A. 1998; 95(18): 10734-10739. [ https://doi.org/10.1073/pnas.95.18.10734 ].
» https://doi.org/10.1073/pnas.95.18.10734 - 95 Ramboz S , et al . Serotonin receptor 1A knockout: an animal model of anxiety-related disorder. Proc Natl Acad Sci U S A. 1998; 95(24): 14476-14481. [ https://doi.org/10.1073/pnas.95.24.14476 ].
» https://doi.org/10.1073/pnas.95.24.14476 - 96 Martin M, Ledent C, Parmentier M, Maldonado R, Valverde O. Involvement of CB1 cannabinoid receptors in emotional behaviour. Psychopharmacol. 2002; 159(4): 379-387. [ https://doi.org/10.1007/s00213-001-0946-5 ].
» https://doi.org/10.1007/s00213-001-0946-5 - 97 Degroot A, Nomikos GG. Genetic deletion and pharmacological blockade of CB1 receptors modulates anxiety in the shock-probe burying test. Eur J Neurosci. 2004; 20(4): 1059-1064. [ https://doi.org/10.1111/j.1460-9568.2004.03556.x ].
» https://doi.org/10.1111/j.1460-9568.2004.03556.x - 98 Witkin JM , Tzavara ET , Nomikos GG . A role for cannabinoid CB1 receptors in mood and anxiety disorders. Behav Pharmacol. 2005; 16(5-6): 315-331. [ https://doi.org/10.1097/00008877-200509000-00005 ].
» https://doi.org/10.1097/00008877-200509000-00005 - 99 Bertoglio LJ , Joca SR , Guimarães FS . Further evidence that anxiety and memory are regionally dissociated within the hippocampus. Behav Brain Res. 2006; 175(1): 183-188. [ https://doi.org/10.1016/j.bbr.2006.08.021 ].
» https://doi.org/10.1016/j.bbr.2006.08.021 - 100 Canteras NS, Resstel LB, Bertoglio LJ, Carobrez AP, Guimarães FS. Neuroanatomy of anxiety. Curr Top Behav Neurosci. 2010; 2: 77-96. [ https://doi.org/10.1007/7854_2009_7 ].
» https://doi.org/10.1007/7854_2009_7 - 101 Steimer T. Animal models of anxiety disorders in rats and mice: some conceptual issues. Dialogues Clin Neurosci. 2011; 13(4): 495-506. [ https://doi.org/10.31887/DCNS.2011.13.4/tsteimer ].
» https://doi.org/10.31887/DCNS.2011.13.4/tsteimer - 102 Giustino TF , Maren S . The role of the medial prefrontal cortex in the conditioning and extinction of fear. Front Behav Neurosci. 2015; 9: 298. [ https://doi.org/10.3389/fnbeh.2015.00298 ].
» https://doi.org/10.3389/fnbeh.2015.00298 - 103 Diehl MM , Moscarello JM , Trask S . Behavioral outputs and overlapping circuits between conditional fear and active avoidance. Neurobiol Learn Mem. 2024; 213: 107943. [ https://doi.org/10.1016/j.nlm.2024.107943 ].
» https://doi.org/10.1016/j.nlm.2024.107943 - 104 Hernandes PM , et al . Sex and estrous cycle-linked differences in the effect of cannabidiol on panic-like responding in rats and mice. Behav Brain Res. 2023; 455: 114663. [ https://doi.org/10.1016/j.bbr.2023.114663 ].
» https://doi.org/10.1016/j.bbr.2023.114663 - 105 Fabris D, et al. Sex-dependent differences in the anxiolytic-like effect of cannabidiol in the elevated plus-maze. J Psychopharmacol. 2022; 36(12): 1371-1383. [ https://doi.org/10.1177/02698811221125440 ].
» https://doi.org/10.1177/02698811221125440 - 106 Chesworth R , Cheng D , Staub C , Karl T . Effect of long-term cannabidiol on learning and anxiety in a female Alzheimer's disease mouse model. Front Pharmacol. 2022; 13: 931384. [ https://doi.org/10.3389/fphar.2022.931384 ].
» https://doi.org/10.3389/fphar.2022.931384 - 107 Yadav-Samudrala BJ, et al. Effects of acute cannabidiol on behavior and the endocannabinoid system in HIV-1 Tat transgenic female and male mice. Frontiers in neuroscience. 2024; 18: 1358555. [ https://doi.org/10.3389/fnins.2024.1358555 ].
» https://doi.org/10.3389/fnins.2024.1358555 - 108 Huffstetler CM, et al. A. Single cannabidiol administration affects anxiety-, obsessive compulsive-, object memory-, and attention-like behaviors in mice in a sex and concentration dependent manner. Pharmacol Biochem Behav. 2023; 222: 173498. [https://doi.org/10.1016/j.pbb.2022.173498].
» https://doi.org/10.1016/j.pbb.2022.173498 - 109 Guimarães FS, Chiaretti TM, Graeff FG, Zuardi AW. Antianxiety effect of cannabidiol in the elevated plus-maze. Psychopharmacol. 1990; 100(4): 558-559. [ https://doi.org/10.1007/BF02244012 ].
» https://doi.org/10.1007/BF02244012 - 110 Resstel LB, Joca SR, Moreira FA, Corrêa FM, Guimarães FS. Effects of cannabidiol and diazepam on behavioral and cardiovascular responses induced by contextual conditioned fear in rats. Behav Brain Res. 2006; 172(2): 294-298. [ https://doi.org/10.1016/j.bbr.2006.05.016 ].
» https://doi.org/10.1016/j.bbr.2006.05.016 - 111 Uribe-Mariño A, et al. Anti-aversive effects of cannabidiol on innate fear-induced behaviors evoked by an ethological model of panic attacks based on a prey vs the wild snake Epicrates cenchria crassus confrontation paradigm. Neuropsychopharmacol. 2012; 37(2): 412-421. [ https://doi.org/10.1038/npp.2011.188 ].
» https://doi.org/10.1038/npp.2011.188 - 112 O'Brien LD, et al. Effect of chronic exposure to rimonabant and phytocannabinoids on anxiety-like behavior and saccharin palatability. Pharmac, Biochem Behav. 2013; 103(3): 597-602. [ https://doi.org/10.1016/j.pbb.2012.10.008 ].
» https://doi.org/10.1016/j.pbb.2012.10.008 - 113 Almeida V, et al. Cannabidiol exhibits anxiolytic but not antipsychotic property evaluated in the social interaction test. Prog Neuropsychopharmacol Biol Psychiatry. 2013; 41: 30-35. [ https://doi.org/10.1016/j.pnpbp.2012.10.024 ].
» https://doi.org/10.1016/j.pnpbp.2012.10.024 - 114 Cheng D, Low JK, Logge W, Garner B, Karl T. Chronic cannabidiol treatment improves social and object recognition in double transgenic APPswe/PS1?E9 mice. Psychopharmacology. 2014; 231(15): 3009-3017. https://doi.org/10.1007/s00213-014-3478-5 ].
» https://doi.org/10.1007/s00213-014-3478-5 - 115 Nardo M, Casarotto PC, Gomes FV, Guimarães FS. Cannabidiol reverses the mCPP-induced increase in marble-burying behavior. Fundam Clin Pharmacol. 2014 Oct; 28(5): 544-50. [ https://doi.org/10.1111/fcp.12051 ].
» https://doi.org/10.1111/fcp.12051 - 116 Schiavon AP , Bonato JM , Milani H , Guimarães FS , Oliveira RMW . Influence of single and repeated cannabidiol administration on emotional behavior and markers of cell proliferation and neurogenesis in non-stressed mice. Prog Neuropsychopharmacol Biol Psychiatry. 2016; 64: 27-34. [ https://doi.org/10.1016/j.pnpbp.2015.06.017 ].
» https://doi.org/10.1016/j.pnpbp.2015.06.017 - 117 Zieba J , et al . Cannabidiol (CBD) reduces anxiety-related behavior in mice via an FMRP-independent mechanism. Pharmacol Biochem Behav. 2019; 181: 93-10. [ https://doi.org/10.1016/j.pbb.2019.05.002 ].
» https://doi.org/10.1016/j.pbb.2019.05.002 - 118 Chaves YC, et al. Two-weeks treatment with cannabidiol improves biophysical and behavioral deficits associated with experimental type-1 diabetes. Neurosci Lett. 2020; 729: 135020. [ https://doi.org/10.1016/j.neulet.2020.135020 ].
» https://doi.org/10.1016/j.neulet.2020.135020 - 119 Assareh N , Gururajan A , Zhou C , Luo JL , Kevin RC & Arnold JC . Cannabidiol disrupts conditioned fear expression and cannabidiolic acid reduces trauma-induced anxiety-related behaviour in mice. Behav Pharmacol. 2020; 31(6): 591-596. [ https://doi.org/10.1097/FBP.0000000000000565 ].
» https://doi.org/10.1097/FBP.0000000000000565 - 120 Gasparyan A , Navarrete F , Manzanares J . Cannabidiol and Sertraline Regulate Behavioral and Brain Gene Expression Alterations in an Animal Model of PTSD. Front Pharmacol. 2021; 12: 694510. [ https://doi.org/10.3389/fphar.2021.694510 ].
» https://doi.org/10.3389/fphar.2021.694510 - 121 Navarrete F, Aracil-Fernández A, Manzanares J. Cannabidiol regulates behavioural alterations and gene expression changes induced by spontaneous cannabinoid withdrawal. Brit J Pharmacol. 2018; 175(13): 2676-2688. [ https://doi.org/10.1111/bph.14226 ].
» https://doi.org/10.1111/bph.14226 - 122 Alegre-Zurano L , López-Arnau R , Luján MÁ , Camarasa J , Valverde O . Cannabidiol Modulates the Motivational and Anxiety-Like Effects of 3,4-Methylenedioxypyrovalerone (MDPV) in Mice. Int J Mol Sci. 2021; 22(15): 8304. [ https://doi.org/10.3390/ijms22158304 ].
» https://doi.org/10.3390/ijms22158304 - 123 Marçal AP, Soares N, Asth L, Moreira FA, Ferreira AVM, Aguiar DC. Cannabidiol ameliorates the anxiogenic and compulsive-like behaviors induced by chronic consumption of a high-carbohydrate diet in male mice. Metab Brain Dis. 2022; 37(8): 2711-2718. [ https://doi.org/10.1007/s11011-022-01071-9 ].
» https://doi.org/10.1007/s11011-022-01071-9 - 124 Jinks AL, McGregor IS. Modulation of anxiety-related behaviours following lesions of the prelimbic or infralimbic cortex in the rat. Brain Res. 1997 Oct 24; 772(1-2): 181-90. [ https://doi.org/10.1016/s0006-8993 (97)00810-x].
» https://doi.org/10.1016/s0006-8993 (97)00810-x - 125 Sotres-Bayon F, Quirk GJ. Prefrontal control of fear: more than just extinction. Curr Opin Neurobiol. 2010 Apr; 20(2): 231-5. [ https://doi.org/10.1016/j.conb.2010.02.005 ].
» https://doi.org/10.1016/j.conb.2010.02.005 - 126 Albert PR , Vahid-Ansari F , Luckhart C . Serotonin-prefrontal cortical circuitry in anxiety and depression phenotypes: pivotal role of pre- and post-synaptic 5-HT1A receptor expression. Front Behav Neurosci. 2014 Jun 6; 8: 199. [ https://doi.org/10.3389/fnbeh.2014.00199 ].
» https://doi.org/10.3389/fnbeh.2014.00199 - 127 Santana N, Bortolozzi A, Serrats J, Mengod G, Artigas F. Expression of serotonin1A and serotonin2A receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb Cortex. 2004 Oct; 14(10): 1100-9. [ https://doi.org/10.1093/cercor/bhh070 ].
» https://doi.org/10.1093/cercor/bhh070 - 128 Yuen EY , Jiang Q , Chen P , Feng J , Yan Z . Activation of 5-HT2A/C receptors counteracts 5-HT1A regulation of n-methyl-D-aspartate receptor channels in pyramidal neurons of prefrontal cortex. J Biol Chem. 2008 Jun 20; 283(25): 17194-204. [ https://doi.org/10.1074/jbc.M801713200 ].
» https://doi.org/10.1074/jbc.M801713200 - 129 Lladó-Pelfort L, Santana N, Ghisi V, Artigas F, Celada P. 5-HT1A receptor agonists enhance pyramidal cell firing in prefrontal cortex through a preferential action on GABA interneurons. Cereb Cortex. 2012 Jul; 22(7): 1487-97. [ https://doi.org/10.1093/cercor/bhr220 ].
» https://doi.org/10.1093/cercor/bhr220 - 130 Celada P , Puig MV , Artigas F . Serotonin modulation of cortical neurons and networks. Front Integr Neurosci. 2013 Apr 19; 7: 25. [ https://doi.org/10.3389/fnint.2013.00025 ].
» https://doi.org/10.3389/fnint.2013.00025 - 131 Lopez J , Gamache K , Schneider R , Nader K . Memory retrieval requires ongoing protein synthesis and NMDA receptor activity-mediated AMPA receptor trafficking. J Neurosci. 2015 Feb 11; 35(6): 2465-75. [ https://doi.org/10.1523/JNEUROSCI.0735-14.2015 ].
» https://doi.org/10.1523/JNEUROSCI.0735-14.2015 - 132 Wissink S , Meijer O , Pearce D , van Der Burg B , van Der Saag PT . Regulation of the rat serotonin-1A receptor gene by corticosteroids. J Biol Chem. 2000 Jan 14; 275(2): 1321-6. [ https://doi.org/10.1074/jbc.275.2.1321 ].
» https://doi.org/10.1074/jbc.275.2.1321 - 133 Zhuang X, Gross C, Santarelli L, Compan V, Trillat AC, Hen R. Altered emotional states in knockout mice lacking 5-HT1A or 5-HT1B receptors. Neuropsychopharmacol. 1999 Aug; 21(2 Suppl): 52S-60S. [ https://doi.org/10.1016/S0893-133X (99)00047-0].
» https://doi.org/10.1016/S0893-133X (99)00047-0 - 134 Rioja J et al. 5-HT1A receptor activation before acute stress counteracted the induced long-term behavioral effects. Ann N Y Acad Sci. 2004 Jun; 1018: 333-8. [ https://doi.org/10.1196/annals.1296.041 ].
» https://doi.org/10.1196/annals.1296.041 - 135 Puglisi-Allegra S, Andolina D. Serotonin and stress coping. Behav Brain Res. 2015 Jan 15; 277: 58-67. [ https://doi.org/10.1016/j.bbr.2014.07.052 ].
» https://doi.org/10.1016/j.bbr.2014.07.052 - 136 Bickle JG, et al. 5-HT1A Receptors on Dentate Gyrus Granule Cells Confer Stress Resilience. Biol Psychiatry. 2024 Apr 15; 95(8): 800-809. [ https://doi.org/10.1016/j.biopsych.2023 ].
» https://doi.org/10.1016/j.biopsych.2023 - 137 Morena M , Leitl KD , Vecchiarelli HA , Gray JM , Campolongo P , Hill MN . Emotional arousal state influences the ability of amygdalar endocannabinoid signaling to modulate anxiety. Neuropharmacology. 2016 Dec; 111: 59-69. [ https://doi.org/10.1016/j.neuropharm.2016.08.020 ].
» https://doi.org/10.1016/j.neuropharm.2016.08.020 - 138 Linge R et al. Cannabidiol induces rapid-acting antidepressant-like effects and enhances cortical 5-HT/glutamate neurotransmission: role of 5-HT1A receptors. Neuropharmacol. 2016; 103: 16-26. [ https://doi.org/10.1016/j.neuropharm.2015.12.017 ].
» https://doi.org/10.1016/j.neuropharm.2015.12.017 - 139 Abame MA et al . Chronic administration of synthetic cannabidiol induces antidepressant effects involving modulation of serotonin and noradrenaline levels in the hippocampus. Neurosci Lett. 2021 Jan 23; 744: 135594. [ https://doi.org/10.1016/j.neulet.2020.135594 ].
» https://doi.org/10.1016/j.neulet.2020.135594 - 140 Guldager MB , Biojone C , da Silva NR , Godoy LD , Joca S . New insights into the involvement of serotonin and BDNF-TrkB signalling in cannabidiol's antidepressant effect. Prog Neuropsychopharmacol Biol Psychiatry. 2024 Jul 13; 133: 111029. [ https://doi.org/10.1016/j.pnpbp.2024.111029 ].
» https://doi.org/10.1016/j.pnpbp.2024.111029 - 141 Mitchell JR, et al. Darting across space and time: parametric modulators of sex-biased conditioned fear responses. Learn Mem. 2022 Jun 16; 29(7): 171-180. [ https://doi.org/10.1101/lm.053587.122 ].
» https://doi.org/10.1101/lm.053587.122 - 142 Mitchell JR, et al. Behavioral and neural correlates of diverse conditioned fear responses in male and female rats. bioRxiv [Preprint]. 2024 Aug 21; 2024.08.20.608817. [ https://doi.org/10.1101/2024.08.20.608817 ].
» https://doi.org/10.1101/2024.08.20.608817 - 143 Werle I , Bertoglio LJ . Psychedelics: A review of their effects on recalled aversive memories and fear/anxiety expression in rodents. Neurosci Biobehav Rev. 2024 Dez; 167: 105899. [ https://doi.org/10.1016/j.neubiorev.2024.105899 ].
» https://doi.org/10.1016/j.neubiorev.2024.105899 - 144 Han K , Wang JY , Wang PY , Peng YC . Therapeutic potential of cannabidiol (CBD) in anxiety disorders: A systematic review and meta-analysis. Psychiatry Res. 2024 Sep; 339: 116049. [ https://doi.org/10.1016/j.psychres.2024.116049 ].
» https://doi.org/10.1016/j.psychres.2024.116049 - 145 Crippa JA, et al. Effects of cannabidiol (CBD) on regional cerebral blood flow. Neuropsychopharmacol. 2004 Feb; 29(2): 417-26. [ https://doi.org/10.1038/sj.npp.1300340 ].
» https://doi.org/10.1038/sj.npp.1300340 - 146 Das RK, et al. Cannabidiol enhances consolidation of explicit fear extinction in humans. Psychopharmacology. 2013 Apr; 226(4): 781-92. [ https://doi.org/10.1007/s00213-012-2955-y ].
» https://doi.org/10.1007/s00213-012-2955-y - 147 Bolsoni LM, Crippa JAS, Hallak JEC, Guimarães FS, Zuardi AW. The anxiolytic effect of cannabidiol depends on the nature of the trauma when patients with post-traumatic stress disorder recall their trigger event. Braz J Psychiatry. 2022 May-Jun; 44(3): 298-307. [ https://doi.org/10.1590/1516-4446-2021-2317 ].
» https://doi.org/10.1590/1516-4446-2021-2317 - 148 Raut SB , et al . Diverse therapeutic developments for post-traumatic stress disorder (PTSD) indicate common mechanisms of memory modulation. Pharmacol Ther. 2022 Nov; 239: 108195. [ https://doi.org/10.1016/j.pharmthera.2022.108195 ].
» https://doi.org/10.1016/j.pharmthera.2022.108195 - 149 Kindt M , Elsey JWB . A paradigm shift in the treatment of emotional memory disorders: Lessons from basic science. Brain Res Bull. 2023 Jan; 192: 168-174. [ https://doi.org/10.1016/j.brainresbull.2022.11.019 ].
» https://doi.org/10.1016/j.brainresbull.2022.11.019 - 150 Kida S. Reconsolidation/destabilization, extinction and forgetting of fear memory as therapeutic targets for PTSD. Psychopharmacol. 2019 Jan; 236(1): 49-57. [ https://doi.org/10.1007/s00213-018-5086-2 ].
» https://doi.org/10.1007/s00213-018-5086-2 - 151 Lookfong NA, Raup-Konsavage WM, Silberman Y. Potential Utility of Cannabidiol in Stress-Related Disorders. Cannabis Cannabinoid Res. 2023 Apr; 8(2): 230-240. [ https://doi.org/10.1089/can.2022.0130 ].
» https://doi.org/10.1089/can.2022.0130 - 152 Luján MÁ , Valverde O . The Pro-neurogenic Effects of Cannabidiol and Its Potential Therapeutic Implications in Psychiatric Disorders. Front Behav Neurosci. 2020 Jun 26; 14: 109. [ https://doi.org/10.3389/fnbeh.2020.00109 ].
» https://doi.org/10.3389/fnbeh.2020.00109 - 153 Domingos LB, Silva NR, Chaves Filho AJM, Sales AJ, Starnawska A, Joca S. Regulation of DNA Methylation by Cannabidiol and Its Implications for Psychiatry: New Insights from In Vivo and In Silico Models. Genes. 2022 Nov 20; 13(11): 2165. [ https://doi.org/10.3390/genes13112165 ].
» https://doi.org/10.3390/genes13112165 - 154 Kim J , Choi H , Kang EK , Ji GY , Kim Y , Choi IS . In Vitro Studies on Therapeutic Effects of Cannabidiol in Neural Cells: Neurons, Glia, and Neural Stem Cells. Molecules. 2021 Oct 8; 26(19): 6077. [ https://doi.org/10.3390/molecules26196077 ].
» https://doi.org/10.3390/molecules26196077 - 155 Hartmann A , Vila-Verde C , Guimarães FS , Joca SR , Lisboa SF . The NLRP3 Inflammasome in Stress Response: Another Target for the Promiscuous Cannabidiol. Curr Neuropharmacol. 2023; 21: 284-308. [ https://doi.org/10.2174/1570159X20666220411101217 ].
» https://doi.org/10.2174/1570159X20666220411101217 - 156 Ling T , et al . Purinergic Astrocyte Signaling Driven by TNF-a After Cannabidiol Administration Restores Normal Synaptic Remodeling Following Traumatic Brain Injury. Neuroscience. 2024 May 3; 545: 31-46. [ https://doi.org/10.1016/j.neuroscience.2024.03.002 ].
» https://doi.org/10.1016/j.neuroscience.2024.03.002 - 157 Gomes FV, Llorente R, Del Bel EA, Viveros MP, López-Gallardo M, Guimarães FS. Decreased glial reactivity could be involved in the antipsychotic-like effect of cannabidiol. Schizophr Res. 2015 May;164(1-3):155-63. [ https://doi.org/10.1016/j.schres.2015.01.015 ].
» https://doi.org/10.1016/j.schres.2015.01.015 - 158 Ye H , Wan Y , Wang X , Wang S , Zhao X , Wang X , et al . Cannabidiol Protects Against Neurotoxic Reactive Astrocytes-Induced Neuronal Death in Mouse Model of Epilepsy. J Neurochem. 2025 Mar; 169(3): e70038. [ https://doi.org/10.1111/jnc.70038 ].
» https://doi.org/10.1111/jnc.70038 - 159 Kamsrijai U, Charoensup R, Jaidee W, Hawiset T, Thaweethee-Sukjai B, Praman S. Cannabidiol/cannabidiolic acid-rich hemp (Cannabis sativa L.) extract attenuates cognitive impairments and glial activations in rats exposed to chronic stress. J Ethnopharmacol. 2025 Feb 10; 338(Pt 3): 119113. [ https://doi.org/10.1016/j.jep.2024.119113 ].
» https://doi.org/10.1016/j.jep.2024.119113 - 160 Xie G , Gao X , Guo Q , Liang H , Yao L , Li W , et al . Cannabidiol ameliorates PTSD-like symptoms by inhibiting neuroinflammation through its action on CB2 receptors in the brain of male mice. Brain Behav Immun. 2024 Jul; 119: 945-964. [ https://doi.org/10.1016/j.bbi.2024.05.016 ].
» https://doi.org/10.1016/j.bbi.2024.05.016
-
Fontes de financiamento:
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; 304851/2022-1; 140296/2023-9); Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES; 88887.953385/2024-00); Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina (FAPESC; 20/2024; 733/2024).
Datas de Publicação
- Publicação nesta coleção
26 Maio 2025 - Data do Fascículo
Maio 2025
Histórico
- Recebido
17 Dez 2024 - Aceito
09 Abr 2025

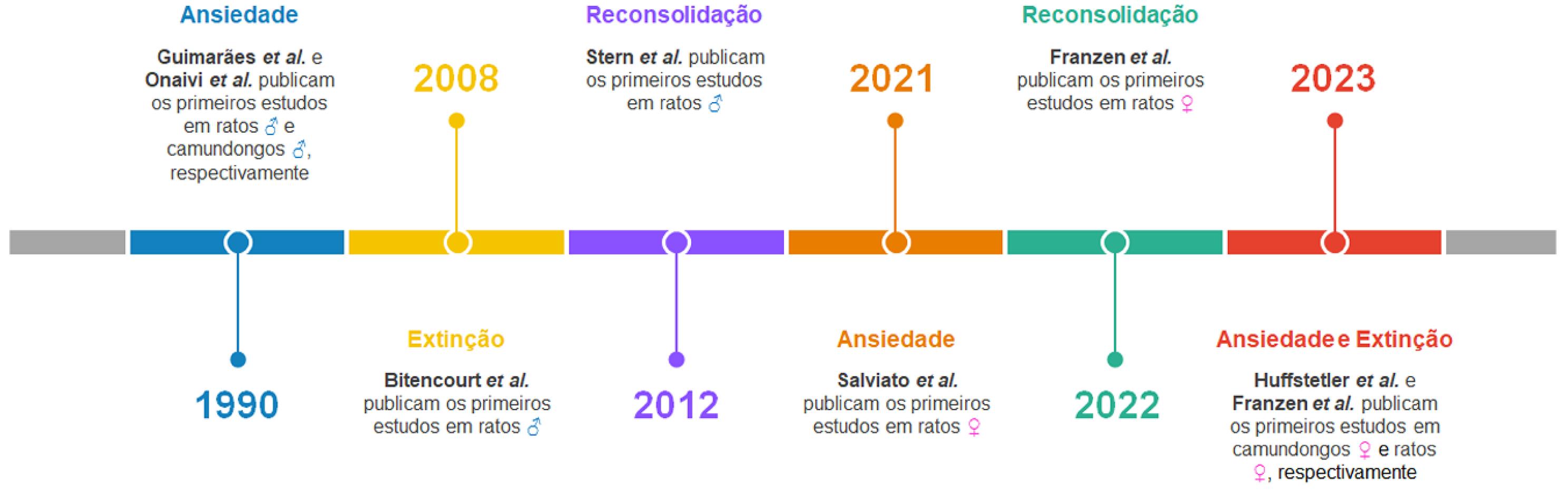
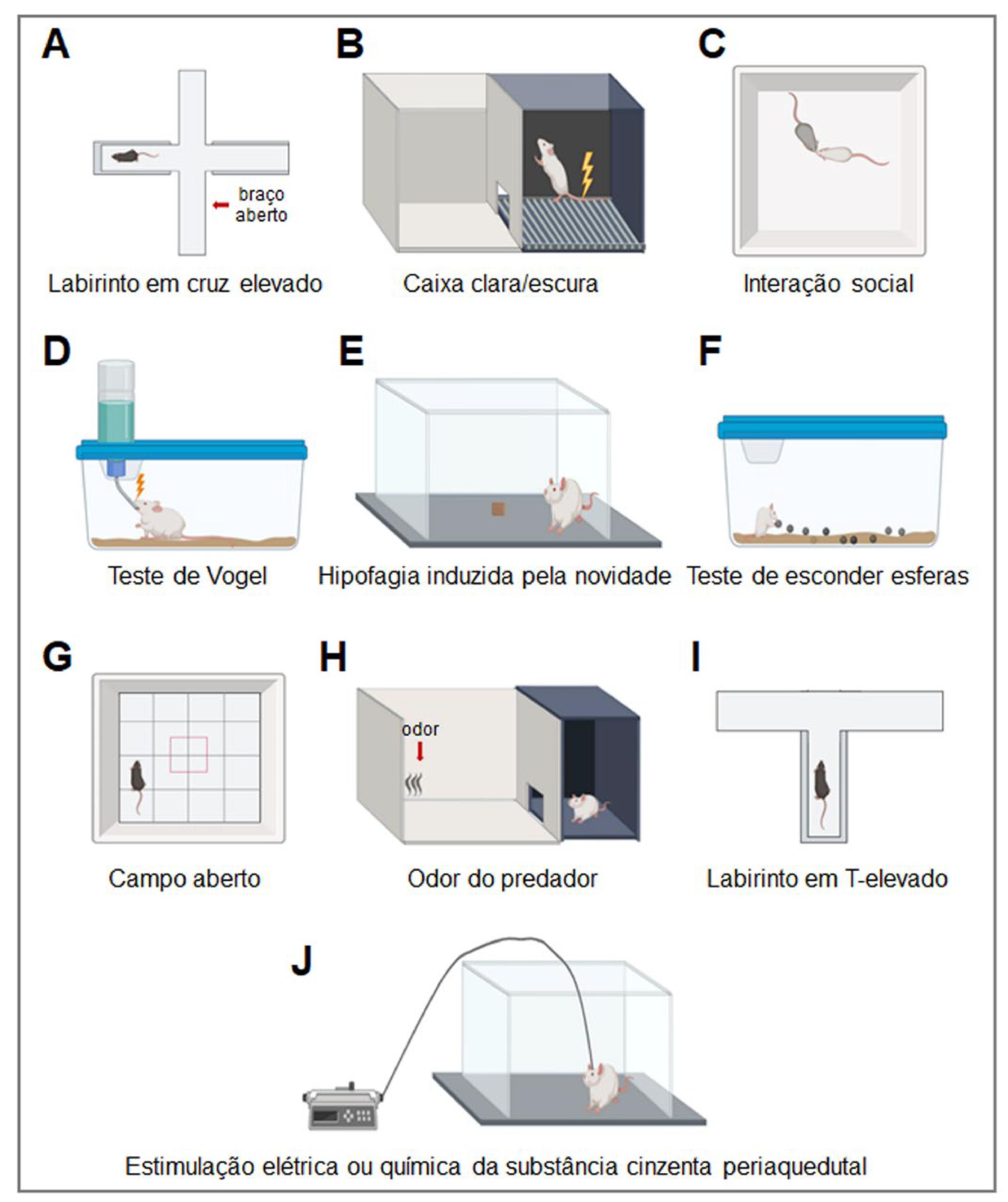 Fonte: Criada com BioRender.com
Fonte: Criada com BioRender.com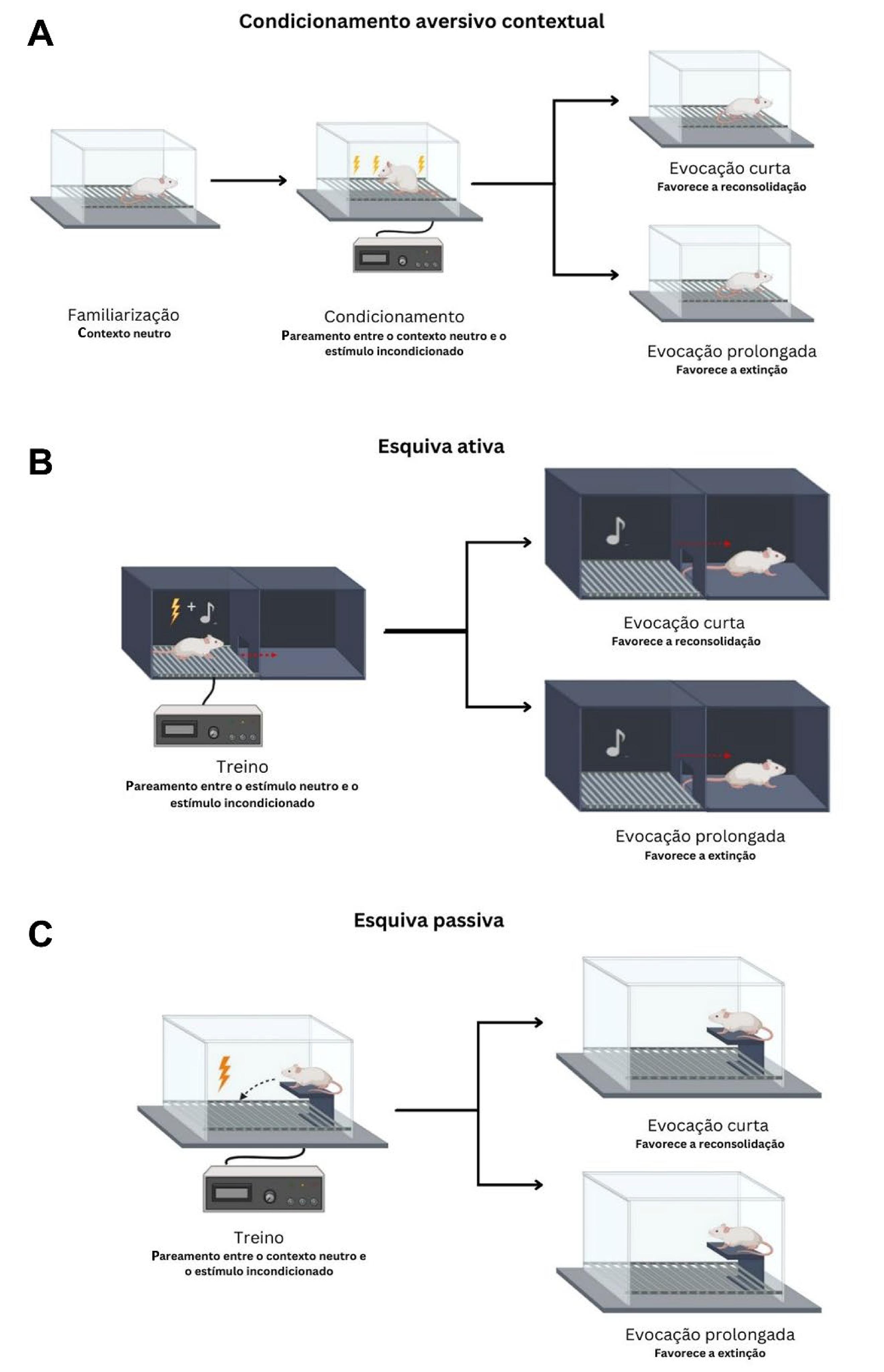 Fonte: Criada com BioRender.com
Fonte: Criada com BioRender.com