Revisão
Contributions of bioinformatics to study natural antifungals: review in a pharmacological context
Abstract
In the last decades, computational techniques have become valuable tools for research of biological systems. They are applied to biochemistry, immunology, pharmacology, etc. Regarding antimicrobial agents, the search for new medicaments or pharmacological strategies is fully justified because of different reasons such as emergence of resistance and drug interactions, particularly in immunocompromised patients. In this work we proposed to update knowledge about application of computational methods to research on natural antifungal compounds. We searched scientific findings through electronic databases according to pertinence and relevance of the information. Then, we analyzed and selected articles in order to organize the information in different items according to set schedules. We updated information about in silico studies regarding new and already known natural antifungal agents. Data were then placed in a pharmacological context regarding molecular targets, mechanisms of therapeutic and toxic effects, as well as resistance mechanisms and their reversion, with special focus on human mycoses. Thus, in this review we summarized important contributions of bioinformatics studies to the design and discovery of new potential agents. We also integrated data related to pharmacology and other disciplines in order to provide a framework for the bioinformatics findings.
- Keywords:
- Bioinformatics.
- Computational studies.
- Theoretical studies.
- Antifungal agents.
- Natural products.
Introduction
Computational techniques have become valuable tools for drug studies over the last decades. When they are applied to biochemical phenomena, bioinformatics arises. It is defined as the science of managing and analyzing biological data using advanced computing techniques, according to the International Human Genome Sequencing Consortium[1].
According to other authors, bioinformatics would be a practical discipline that employs a wide range of computational techniques, database design and data mining, macromolecular geometry, phylogenetic tree construction, prediction of protein structure and function, gene finding and expression data clustering. Some authors have integrated a variety of computational methods and heterogeneous data sources[2].
Bioinformatics allows the knowledge of new chemical structures and functions and new targets to drive drug design in a more precise and accurate way. Since 1980, the idea of rational use of drugs arises applying the "rational design of new drugs" (structure-guided drug design) and using technologies related to structural biology to generate a target protein and small molecules in silico as basis for drug generation[3] (FIGURE 1).

Computational methods offer the advantage of low costs and rapid acquisition of information. They could be complemented by experimental research to optimize the route of scientific work. In this way, bioinformatics serves to expand the horizon of the experimental conclusions, and vice versa. It is being applied to different fields as biochemistry, biophysics, genetics, pharmacology, toxicology, immunology, bromatology and agriculture. Bioinformatics is largely involved in the development of new drugs. In 2016, Tamay-Cach et al.[4] summarized the methods most used in the search and design of drugs used against bacteria, virus and parasites.
Our labs have complemented experimental and computational studies on antifungal agents and updated contributions of bioinformatics to pharmacology of natural products [5-9]. These investigations take place in the frame of recommendations of the World Health organization (WHO), which promotes the search of pharmacological products from natural resources. Considering current increase in resistance to antimicrobial agents, particularly to antifungal ones (as discussed below), the discovery of natural drugs and biological activities is absolutely necessary for the safe treatment of infections.
In this review we update information on advances in natural antifungal agents, particularly regarding bioinformatics studies. We integrate data related to pharmacology and other disciplines in order to provide a framework for the bioinformatics findings.
Thus, this review has been prepared by collecting information about natural antifungal products with therapeutic efficacy, their molecular targets and the eventual resistance developed. We have looked especially for bioinformatics data related to the study of the items above mentioned.
Material and methods
Methodology used for the review
Information about in silico studies regarding new and already known natural antifungal agents was updated. Scientific findings were searched and screened through electronic databases according to article titles and abstract text. Selected articles were then analyzed in order to organize the information in different items according to set schedules. Data were related to molecular targets, mechanisms of therapeutic and toxic actions as well as resistance mechanisms and their reversion, with special focus on human mycoses.
Electronic databases used: PubMed (Medline), Scopus, Lilacs, Web of Science, Google Scholar.
The terms used in the searches were related to the above mentioned data and may be summarized as follows: in silico, bioinformatics, computational, theoretical, docking studies AND/OR natural antifungal, antimicrobial products, antifungal therapeutic and adverse effects, resistance mechanisms, bioinformatics studies and biological activities related to human mycoses.
Criteria for inclusion or exclusion of articles were the scientific pertinence and relevance of the information reported in relation to the topics focused.
Studies on natural products
Different biological activities
Since prehistoric times, people have used natural resources for nutritional and medicinal purposes. For example, they consume numerous plants for the preparation of meals and beverages using different techniques that may alter the chemical composition of some compounds. Plants can be used as cheap sources of different nutritive substances like proteins, carbohydrates, lipids and crude fibers. Numerous studies about nutritional quality of diverse plants have been made[10].
In recent years, the interest in the development of functional foods and nutraceuticals has increased due to the beneficial health effects associated with their intake. Since 1950, microalgae were mainly viewed as an alternative food source and functional biomolecules[11].
Custódio et al.[12] found that organic and water extracts of some microalgae species exhibit antioxidant, antitumoral activities and acetyl cholinesterase inhibition, besides a highly unsaturated fatty acids profile. These results suggest that those microalgae, particularly I. galbana T-ISO, could be a source of biomolecules for pharmaceutical and food industries.
Regarding medicinal fields, plants belonging to diverse families are popularly used to treat different diseases. Some scientific works have identified various compounds which are responsible for many therapeutic effects[13,14].
The Argentine folk medicine employs many plants to counteract diverse diseases such as catarrh, bronchitis, pneumonia and diarrheas. Our labs have carried out several studies on antimicrobial activity related to some of the mentioned plants. The antimicrobials reported can be extracted easily in an aqueous medium. This observation may be useful in isolating these principles from consumption residues (i.e. fruit pericarp of oranges and pomegranates) at a reduced cost. Moreover, Citrus aurantium L. (bitter orange) pericarps are codified in the Pharmacopeias of Argentina and Great Britain. They contain flavonoids as naringin and hesperidin, which are active against Candida albicans and dermatophytes. Besides antimicrobial properties, flavonoids show antioxidant anti-inflammatory, antiasmathic, antiagregant and hormonal effects[15-17].
We have conducted other studies in relation to natural compounds isolated from plants of Argentina. The compounds studied were mainly flavonoids with different chemical formulae and some of them were present in essential oils. Others have an infrequent distribution in nature and exhibit specific biological activities that allow them to be classified as potential medicaments to collaborate with others existing in the pharmaceutical market[5,18,19]. Other compounds showing bioactivity are alkaloids, lectins, saponins and peptides, which were found in plants as well as in microorganism's cultures contained in soil samples[20].
β -citronellol, a monoterpenoid contained in several plant essential oils, modulates expression of ERG genes and ergosterol levels as part of its antifungal effect[21]. The results suggest that β-citronellol has various modes of action, including inhibition of morphological transition in C. albicans and decrease of the secretion of hydrolytic enzymes involved in the early stage of infection as well as modulation of the expression of associated genes. In fact, it has a significant inhibitory effect on biofilm formation and secretion of extracellular proteinases and phospholipases as well in the membrane ergosterol levels.
Among the medicaments of vegetal origin, we can mention the cardiotonic digoxin, the analgesic morphine, the antitumoral vincristine and the antiseptic agent eugenol, widely used in Odontology. Coming from other natural resources, the betalactamyc antibiotics are produced by fungal cultures and the polypeptides ones by soil bacteria.
Recently, investigation on natural resources has been increased due to vital need for new agents. In this sense, it can be also applied to different fields such as medicine, pharmacy, biochemistry, bromatology, cosmetology, agriculture, etc.
Research on natural antifungal agents. Human mycoses
Fungal infections are a public health problem. The most common fungal pathogens for humans are the species of Candida and Aspergillus. Opportunistic yeast Candida, mainly C. albicans, is responsible of 50-90% human candidiasis[22].
The use of antimicrobial drugs always carries the risk that resistance appears due to adaptable nature of the microorganism populations[23]. The need for safe and effective antifungal drugs has increased in parallel with the expansion of immunocompromised patients at risk for fungal infections.
The major limitations associated with currently used antifungal compounds are low efficacy rate and severe side effects. Thus, it is necessary to search for new compounds with low toxicity on human cells.
Commonly used antifungal drugs belong to various pharmacological groups and are characterized by different mechanisms of action and spectrum of activity. They act mainly in cell membrane, cell wall and intracellular entities[24]. Several actions have been stated for antifungal therapy; usually they are related to synthesis inhibition of ergosterol, squalene epoxidase, glucan, chitin, nucleic acids, proteins and microtubules[25].
There are several chemical structures which have demonstrated antifungal activity such as those belonging to the groups of azoles, morpholines and other heterocyclic compounds. Different natural compounds have been reported as possible antifungal agents, such as phenols, flavonoids, coumarins, quinones, saponins, xanthones, alkaloids and terpenoids[26,27].
For many years, the treatment of invasive fungal infections was limited to amphotericin B. Afterwards, azoles and echinocandins have been mostly used for the treatment of fungal infections with high mortality rate[28].
Antifungal drug targets
Sterol 14 alpha demethylase CYP51
Sterol 14 alpha demethylase is the main enzyme involved in ergosterol synthesis. Due to its inhibition, depletion of ergosterol is produced and accumulation of 14-methyl sterols results in impaired fungal cell growth. In Candida albicans, the enzyme is called CYP51 and is a member of Cytochrome P450 family that converts lanosterol into 4,4'-dimethylcholesta-8,14,24-triene-3-beta-ol. The essential role of CYP51 enzyme in fungi and yeast metabolism makes it an important target for antifungal drug design. Thus, azoles interfere with the synthesis of ergosterol by inhibiting this enzyme[29,30].
There are many studies that apply informatics tools to natural and synthetic products. In 2015, Irfan and Abid[31] carried out docking studies to build a three dimensional model of Candida albicans CYP51 and study its interactions with eighteen newly synthesized triazoles derived from naturally bioactive scaffolds.
Docking studies using I-TASSER (iterative threading assembly refinement) server for 3D showed that a compound containing the quinoline ring NT-03 binds with higher affinity the enzyme at the predicted site for medicaments, in accordance with experimental bioactivity data. The authors concluded that similar triazole derivatives can be developed as more effective antifungal agents.
N-Myristoyl transferase
Other therapeutic target is the enzyme N-Myristoyl transferase (NMT), which catalyzes the transfer of the 14-carbon saturated fatty acid myristate from myristoyl-CoA to the N-terminal glycine residue of a variety of eukaryotic cellular and viral proteins. NMT exists as isoenzymes that are involved in a wide variety of biological processes and function as protein kinases, kinase substrates, protein phosphatases, α-subunits of many heterotrimeric G-proteins and endothelial cell nitric oxide synthase[32]. NMT participates in diverse biological processes, including signal transduction cascades and apoptosis. It has even been employed as drug target for human pathogens like Candida sp and proposed in antitrypanosomatid and antimalarial therapeutics[33,34].
Guerrero-Perilla et al.[35] studied the docking of thirty-two natural compounds to the active site of N-myristoyl transferase (NMT) in comparison with a previously reported synthetic inhibitor. Among the tested compounds, alkaloids, flavonoids, xanthones and quinones exhibited a stronger mean interaction with NMT than terpenoids, coumarins and phenolic derivatives. Particularly, affinities for an aporphine alkaloid, a prenylated flavonoid and two xanthones resulted to be comparable with the reference inhibitor. The results allowed selecting three possible lead structures to continue search for antifungal drugs.
Mevalonate-5-diphosphate decarboxylase and other molecular targets of wortmannin
Fungal strains from soil samples are able to produce narrow to broad spectrum antifungal metabolites. Among them, Penicillium sp have ubiquitous occurrence in diverse ecological niches and demonstrate some important functional attributes associated with plant growth such as solubilisation of phosphorus, biological control of root diseases and phytohormone production[36]. Several metabolites from Penicillium spp, such as paxisterol and compactin, have been reported for analgesic and antifungal properties in animals[37].
Using soil samples as antifungal resources, Singh et al.[38] isolated a cell-permeable metabolite known as wortmannin (Wtmn) from Penicillium radicum. It showed a broad spectrum on antifungal activity and irreversible inhibition of phosphoinositide 3-kinase (PI3K). Among 38 fungal species, wortmannin was active against various strains of Candida sp, Cryptococcus terreus, Trichophyton rubrum, Rhizoctonia oryzae, Aspergillus versicolor, Aspergillus fumigates, Aureobasidium pullulans, Fusarium moniliforme and Saccharomyces cerevisiae.
Virtual screening predicted that the hydrophobic estrogen-related wortmannin may inhibit some targets more efficiently than known antifungal agents such as voriconazole and nikkomycin. The targets are: a) mevalonate-5-diphosphate decarboxylase (1FI4), responsible for sterol/isoprenoid biosynthesis; b) exocyst complex component SEC3 (3A58) where Rho- and phosphoinositide-dependent localization is present and c) Kre2p/ Mnt1p, a Golgi alpha 1,2-mannosyltransferase involved in the biosynthesis of yeast cell wall glycoproteins Singh et al.[38].
The authors suggest that wortmannin produced by P. radicum is a promising lead compound which could be potentially used as efficient antifungal drug in the near future after appropriate structural modifications to reduce toxicity and improve stability.
Thioredoxin reductase on paracoccidioidomycosis
Invasive fungal infections have been important causes of morbidity and mortality, especially in the large population of immunocompromised patients[39]. These infections have also increased significantly and represent a serious public health problem, because they are associated with prolonged length of hospital stay and, consequently, high costs for critically ill patients. The most important microorganisms agents involved in opportunistic mycoses are Candida spp., Cryptococcus neoformans and Aspergillus spp, whereas the most commonly endemic mycoses are due to Histoplasma capsulatum, Paracoccidioides brasiliensis and Coccidioides spp.
P. brasilisiensis is known to cause the systemic granulomatous mycosis paracoccidioidomycosis. However, recent phylogenetic studies have established the involvement of a new species, named Paracoccidioides lutzii[40]. The disease is restricted to Latin America, where ten million people are supposed to be infected with this fungus. The chronic form is the most frequent one among adult males and progresses slowly. The acute form is fast (weeks or months) and more severe, leading to high mortality levels[41].
In this context, one promising fungal target is the thioredoxin reductase (TRR1), a flavoprotein enzyme involved in the oxidative stress resistance, regulation of DNA and methionine biosynthesis, cell growth, gene transcription and apoptosis. Abadioet al.[42] studied the docking of P. lutzii TRR1 to 3,000 small molecules that had showed antifungal activity against Paracoccidiodes spp. Twelve compounds were selected from the results and then tested experimentally on Paracoccidioides lutzii and P. brasiliensis. Three of the compounds tested showed inhibitory activity against the fungal enzyme and were patented. Another compound demonstrated both selectivity and high antifungal activity against Paracoccidioides species. Once again, a rational combination of molecular modeling simulations and virtual screening of new drugs has provided a cost-effective solution to an early-stage medicinal challenge.
Emergence and reversion of antifungal resistance
In the three past decades a significant increase in the prevalence of resistance to antimicrobial agents has been detected. Resistance has important implications for morbidity, mortality and health care in hospitals as well as in the community.
The study of resistance to antifungal agents has lagged behind that of antibacterial resistance for several reasons. Perhaps fungal diseases were not recognized as important pathologies until relatively recent times. For example, the annual death rate due to candidiasis was steady between 1950 and 1970. Since 1970, this rate increased significantly in association with more widespread use of therapies that decrease the immune system, the indiscriminate use of broad-spectrum antibacterial agents, the advent of AIDS, etc.[43].
For more than 30 years, amphotericin B was the sole drug available to control serious fungal infections, in spite of its nephrotoxicity. The approval of azoles in late 1980 meant a major achievement in consideration of the safety/efficacy ratio. However, their widespread use has been correlated with antifungal resistance.
It was tempting to compare mechanism of resistance to antifungal and antibacterial agents. Although it may be useful to develop a perspective common to the two kingdoms, there are some limitations due to their differences in biological structures, the generation time, the pharmacological targets and functions to be inhibited[43].
The mechanisms of resistance to antifungal agents are closely related to those of action. The three main groups of antifungal agents in clinical use, azoles, polyenes and allylamine/thiocarbamates, interact directly or indirectly with ergosterol, which is the predominant component of the fungal cell membrane.
Resistance related to ergosterol synthesis
Resistance linked to this pathway may be mediated either by modification of the enzyme sterol 14 alpha-demethylase or by an overexpression of its encoding gene ERG11 [44]. Some fungal species are naturally resistant to azole drugs due to the chemical structure of the enzyme, which presents polymorphism. For example, Aspergillus fumigates intrinsic resistance is related to the isoenzyme known as Cyp51Ap. Edlind et al.[45] compared in silico the enzyme sequences from Candida albicans and A. fumigatus. They found that the substitution of the polar T315 residue by the non-polar alanine (T315A) in C. albicans enzyme is enough to confer fluconazole resistance to the yeast.
Aminoacid substitutions Y132 and K143 in ERG11 sequences of Candida auris confer a phenotype of resistance to fluconazol similar to that described for Candida albicans[46]. Candida auris, recently recognized MDR yeast found in healthcare settings, is considered a major threat to ICU patients. Worldwide reports of C. auris, which was initially described in 2009, have considerably increased in less than a decade, witnessing its spread in different continents[47,48]. MDR C. auris is causing persistent colonization and invasive infections among hospitalized patients. In 2016, international health agencies including the US CDC[49], ECDC[50] and Pan American Health Organization (PAHO)/WHO[51] warned about it.
Later experimental studies confirmed that the natural polymorphism I301 in the Aspergillus fumigatus enzyme Cyp51Ap is necessary and sufficient to explain the fluconazole intrinsic reduced susceptibility of the yeast. Thus, molecular changes in the enzyme may reverse the resistance to antifungal agents[52].
Aspergillus fumigatus is the most common hyphomycete to cause disease in humans[53,54]. It is intrinsically resistant to ketoconazole and fluconazole but normally susceptible to other available azole antifungal agents as itraconazole and posaconazole[55,56].
Resistance related to efflux pumps
ABC transporters cdr
Polymorphism of other molecules such as drug transporters may also generate antifungal resistance. Efflux pumps are important molecular targets involved in antifungal resistance. Thus, reduced intracellular accumulation of drugs as a consequence of their rapid efflux is another prominent mechanism of resistance in Candida cells[44,57].
Most commonly, genes encoding drug efflux pumps belonging to ATP binding cassette (ATP) and Major Facilitator superfamilies (MFS) of proteins are overexpressed, since ABC and MFS transporters are among the major players that contribute to azole resistance in clinical isolates of Candida.
The genome of C. albicans possesses 28 ABC and 95 MFS proteins; however, only ABC transporters Ca cdr1, Ca cdr2, Cdr6p/Roa1p and MFS transporter Ca mdr1 are known to be multidrug transporters which play major role in drug extrusion from resistant strains. Cdr1 is the major drug exporter in C. albicans and plays a key role in antifungal resistance. Even when Ca cdr2 exhibits 84% identity with Ca cdr1, it has a distinct drug resistance profile[58,59].
ABC proteins of C. albicans are divided into four subfamilies: PDR/ABCG, MDR associated protein (MRP)/ABCC, MDR/ABCB and adrenoleukodystrophy protein/ABCD These transporters are usually located in plasma membrane, specifically Cdr1p, Cdr2p, Cdr3p, and Cdr4p, but can also be found in vacuolar membrane like Adp1[59].
ABC proteins are generally made up of two domains located into the plasmatic membrane and two nucleotide-binding regions situated in the cytoplasm, being the non catalytic nucleotide‐binding site essential in the transport mechanism of Cdr1´s protein[59,60]. The transporters require energy from the ATP hydrolysis which is carried out at the last mentioned sites.
The range of Ca cdr1 substrates varies enormously and includes structurally unrelated compounds such as azoles, lipids and steroids. This promiscuity towards substrates is a characteristic feature of most ABC-type drug transporters and makes their functionality more complex to understand[57]. Nim et al.[61] identified tetrazolium chloride and trimethyltin chloride like new substrates, and also discovered new substrate recognition sites. TMH 1, -2, -4, -8 and -11 harbour multiple overlapping drug-binding sites[59].
Puri et al.[62] made studies to build computational models for classification of substrates and non-substrates of Ca cdr1 and Ca mdr1. Then they selected randomly 67 compounds of different characteristics in order to evaluate toxicity on S. cerevisiae strains over-expressing the mentioned transporters using experimental methods. Berberine, vinblastine and quinidine were found among the natural compounds that result common substrates of Ca cdr1 and Ca mdr1.
In the search for novel modulators or inhibitors of these efflux pump proteins, our labs had reported that the prenylated flavonoid 2',4'-hydroxy - 5'-(1'' '-methylallyl)- 6-prenyl pinocembrine (6PP) inhibits the ATP-dependent cdr transporters through a competitive mechanism and reverses azole resistance in Candida albicans. Besides this, it has an antimicrobial activity by itself [5,6].
In order to further analyze the possible molecular interactions between the transporters cdr1 and cdr2 with the ligands 6PP, fluconazole and adenosine triphosphate (ATP), preliminary bioinformatics studies were conducted. The compound 6PP and fluconazole could compete in different ways by a transmembrane stereo specific site of Candida albicans cdr pumps. The most probable site is shared by cdr1 and cdr2 and contributes selectively with some amino acids. In addition, 6PP may compete with ATP in the cytosolic domain and fluconazole would mimic it [7] (FIGURE 2). Since 6PP chemical formula was reviewed as 2',4'-hydroxy -5'-(1'' '-methylallyl)-8-prenylpinocembrine (8PP), current bioinformatics studies are currently updated.
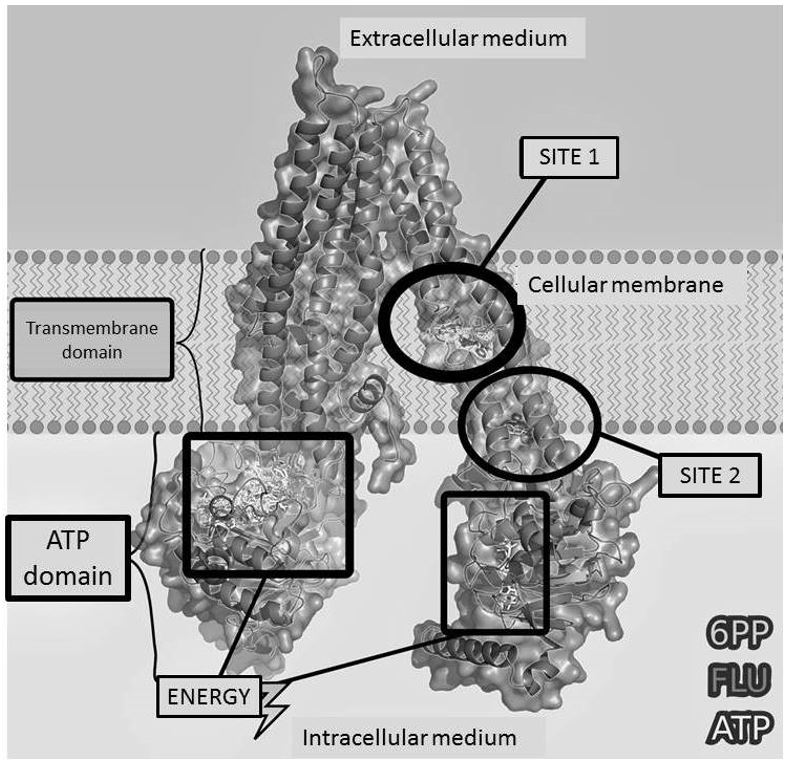
Substitution of residues in Ca cdr1 and other fungal ABC transporters plays an important role in ATP binding and hydrolysis and therefore are essential for drug efflux[57].
ABC transporter pdr5 (Pdr5p)
There is a high structural and functional similarity between Candida albicans cdr1 and Saccharomyces cerevisiae pdr5 (Sc pdr5), even when some distinct functional features tend to distinguish them.
As the best-studied and most abundant ABC transporter in Saccharomyces cerevisiae, pdr5 mediates cellular detoxification and multidrug resistance (MDR). The predicted topological structure of pdr5p shows that it contains two nucleotide binding domains and two transmembrane domains which harbor twelve transmembrane helices, six extracellular loops and four intracellular loops. Pdr5 shares similar secondary topological structure with other similar molecules[63].
To clarify the mechanisms of how pdr5 recognizes and extrudes a wide variety of compounds out of the cells, numerous mutants had been generated by random or site-directed mutagenesis. To investigate the potential loss-of-function mechanism Chen et al.[60] used mutants of the ABC transporter and evaluated its efficacy by comparing results from computational and experimental methods. In order to study the structure–function relationship of pdr5, they used a recently published 3D model of the transporter mutations to perform a study on molecular docking to different compounds.
Mutations adjacent to the end of transmembrane helices 6 and 7 affect drug efflux capacity of yeast pdr5. These mutations might disturb interactions between the drugs and pdr5, preventing the drugs from approaching the intracellular or extracellular portal and subsequently from being exported by the transporter.
Methods used by bioinformatics
Bioinformatics uses different techniques and programs that have been reviewed by Tamay-Cach et al.[4] and Diniz and Canduri[64], in relation to their application in the knowledge of biological systems and in the study of antimicrobial agents. Different bioinformatics approaches discussed by Bencurova et al.[65] help in the identification of novel fungal drug targets.
Among most used methods, virtual screening (VS), docking simulation, molecular dynamics (MD) simulations, biological sequence comparison by alignment, quantitative structure-activity relationship (QSAR), multi-target quantitative structure-activity relationship (mtQSAR) and multi-tasking quantitative structure biological effect relationships (mtk-QSBER) can be mentioned. Some examples and applications are stated below.
Biomolecular docking
According to Chaskar et al.[66], docking methods are concerned with the study of the possible molecular interactions between a ligand and the binding site of a target macromolecule, usually an enzyme, a receptor or a transporter. In this context, Irfan and Abid[31] applied docking methods to study the interactions of diverse triazole drugs with the enzyme lanosterol 14-α demethylase of Candida albicans (see section "Antifungal molecular targets" above).
A new way to find therapeutic targets of a given drug is the inverse docking. In this technique, the ligand is evaluated in silico for binding to different potential targets included into a digital protein cavity database. For example, the natural compound tanshinone has been associated with 10 potential disease targets related to cortisone reductase deficiency and other disorders[67]. Meanwhile, the natural flavanone 8PP could bind to different target macromolecules such as ATP cdr transporters, Hsp90 molecular chaperone and epidermal growth factor receptor[8].
Alignments
Sequential and structural alignment is another bioinformatics method which compares DNA, RNA or protein structures. Study of similarities between compared sequences assigns function to genes and proteins that may be involved not only in medicine but in agriculture and other fields[68,69]. Edlind et al.[45] compared in silico the Cyp51Ap enzyme sequences from Candida albicans and A. fumigatus (see section"Resistance related to ergosterol synthesis" above). The multiple sequence alignment (MSA) is a powerful tool to include a function or a structure by a determined consensus sequence[70].
Conclusion
From its beginning, bioinformatics has grown by leap and bounds. Many laboratory groups in the world perform several bioinformatics studies to have a better vision of the experimental hypothesis or to complement it. Moreover, the use of bioinformatics techniques allows the research groups to direct an experimental design towards a positive outcome, obtaining a virtual preliminary result of what would happen in reality.
Bioinformatics has made important contributions to the study of compounds with diverse pharmacological activities. Among them, antifungal agents deserve special consideration due to the emergence of resistance. In this sense, the search for strategies addressed to reverse it or to find new agents with high efficacy and low toxicity are fully justified, particularly taking into account that other factors, such as drug interactions, may occur. Complementing in silico and experimental methods may be useful to optimize cost of resources and time.
Acknowledgments
This work was granted by the University of Buenos Aires (grant UBACYT 2018, 20020170200079BA). We thank to Milagros Muthular for technical assistance.
References
1. International Human Genome Sequencing Consortium. The Human Genome Project. Information Archive glossary. 2001. [Link].
2. Luscombe NM, Greenbaum D, Gerstein M. What is bioinformatics? A proposed definition and overview of the field. Methods Inf Med. 2001; 40(4): 346-358. ISSN 0026-1270. [PubMed].
3. Peláez F. Paradigmas actuales en las etapas tempranas del proceso de descubrimiento y desarrollo de nuevos fármacos. An Real Soc Española Quim. 2011; 107(1): 36-45. ISSN1575-3417. [Link].
4. Tamay-Cach F, Villa-Tanaca ML, Trujillo-Ferrara JG, Alemán-González-Duhart D, Quintana-Pérez JC, González-Ramírez IA et al. In Silico Studies Most Employed in the Discovery of New Antimicrobial Agents. Curr Med Chem. 2016; 23(29): 3360-3373. ISSN 1875-533X. [CrossRef].
5. Pérez C, Tiraboschi IN, Ortega MG, Agnese AM, Cabrera JL. Further antimicrobial studies on 2´ 4´- dihydroxy- 5´-(1´´´ -dimethylallyl)- 6-prenylpinocembrin from Dalea elegans. Pharm Biol. 2003; 41(3): 171-174, ISSN 1744-5116. [CrossRef].
6. Peralta MA, Calise M, Fornari MC, Ortega MG, Diez RA, Cabrera JL et al. A prenylated flavanone from Dalea elegans inhibits rhodamine 6G efflux and reverses fluconazole resistance in Candida albicans. Pl Med. 2012; 78(10): 981-987. ISSN 0032-0943.
7. Barceló S, Peralta MA, Ortega MG, Cabrera JL, Pérez C. Interacciones moleculares de un flavonoide prenilado con transportadores de antimicóticos dependientes de ATP. Rev Fac Odon UBA. 2014; 29(66): 26-36. ISSN 1668-8538.
8. Barceló S, Farah E, Pérez C. Interacciones de un compuesto de origen vegetal con distintos blancos terapéuticos. In IX international symposium on natural products chemistry and applications. Termas de Chillán, Chile. November 22-25. 2016; 134.
9. Barceló S, Miozza V, Passero P, Farah E, Pérez, C. Aplicaciones de la informática en el estudio de productos naturales. Rev Fac Odontol UBA. 2017; 32(72): 22-31. ISSN 1668-8538. [Link].
10. Saldanha LG, Dwyer JT, Betz JM. Culinary spice plants in dietary supplement products and tested in clinical trials. Adv Nutr. 2016; 7(2): 343-348. ISSN 2156-5376. [CrossRef] [PubMed].
11. ChacónLee TL, GonzálezMaríño GE. Microalgae for "healthy" foods—possibilities and challenge. Comp Rev Food Sci Food Saf. 2010; 9(6): 655–675. ISSN 1541-4337. [CrossRef].
12. Custódio L, Soares F, Pereira H, Barreira L, Vizetto-Duarte C, João Rodrigues M et al. Fatty acid composition and biological activities of Isochrysis galbana TISO, Tetraselmis sp.and Scenedesmus sp.: possible application in the pharmaceutical and functional food industries. J Appl Phycol. 2014; 26(1): 151161. ISSN 1573-5176. [CrossRef].
13. Cicero AFG, Colletti A. Combinations of phytomedicines with different lipid lowering activity for dyslipidemia management: The available clinical data. Phytomedicine. 2016; 23(11): 1113-1118. ISSN 1618-095X. [CrossRef] [PubMed].
14 Oliveira JS, Silva AA, Silva VA Jr. Phytotherapy in reducing glycemic index and testicular oxidative stress resulting from induced diabetes: a review. Braz J Biol. 2017; 77(1): 68-78. ISSN 1678-4375. [CrossRef].
15. Anesini C, Pérez C. Screening of plants used in Argentine folk medicine for antimicrobial activity. J Ethnopharmacol. 1993; 39(2): 119-128. ISSN 0378-8741. [CrossRef].
16. Pérez C, Suarez C. Antifungal activity of plant extracts against Candida albicans. Am J Chin Med. 1997; 25(2): 181-184. ISSN 1793-6853. [CrossRef].
17. Elingold I, Isollabella MP, Casanova M, Celentano AM, Pérez C, Cabrera JL et al. Mitochondrial toxicity and antioxidant activity of a prenylated flavonoid isolated from Dalea elegans. Chem Biol Interact. 2008; 171(3): 294-305. ISSN 1872-7786. [CrossRef] [PubMed].
18. Pérez C, Agnese AM, Cabrera JL. The essential oil of Senecio graveolens (Compositae): Chemical composition and antimicrobial activity tests. J Ethnopharmacol. 1999; 66(1): 91-96. ISSN 0378-8741. [CrossRef] [PubMed].
19. Agnese AM, Pérez C, Cabrera JL. Adesmia aegiceras: antimicrobial activity and chemical study. Phytomedicine. 2001; 8(5): 389-394. ISSN 1618-095X. [CrossRef].
20. Pérez C, Suarez C, Castro G. Production of antimicrobials by Bacillus subtilis MIR 15. J Biotechnol. 1992; 26(2-3): 331-336. ISSN 1873-4863. [CrossRef].
21. Sharma Y, Rastogi SK, Perwez A, Rizvi MA, Manzoor N. β-citronellol alters cell surface properties of Candida albicans to influence pathogenicity related traits. Med Mycol. ISSN 1365-280X. [CrossRef].
22. Vázquez-González D, Perusquía-Ortiz AM, Hundeiker M, Bonifaz A. Opportunistic yeast infections: candidiasis, cryptococcosis, trichosporonosis and geotrichosis. J Dtsch Dermatol Ges. 2013; 11(5): 381-394. ISSN 1610-0387. [CrossRef] [PubMed].
23. Anderson JB. Evolution of antifungal - drug resistance: Mechanisms and pathogen fitness. Nat Rev Microbiol. 2005; 3(7): 547-556. ISSN 1740-1534. [CrossRef].
24. Lewis RE. Current concepts in antifungal pharmacology. Mayo Clin Proc. 2011; 86(8): 805-817. ISSN 1942-5546. [CrossRef] [PubMed].
25. Kathiravan MK, Salake AB, Chothe AS, Dudhe PB, Watode RP, Mukta MS et al. The biology and chemistry of antifungal agents: A review. Bioorg Med Chem. 2012; 20(19): 5678-5698. ISSN 0968-0896. [CrossRef] [PubMed].
26. Arif T, Bhosale JD, Kumar N, Mandal TK, Bendre RS, Lavekar GS et al. Natural products-antifungal agents derived from plants. J Asian NatProd Res. 2009; 11(7): 621-.638. ISSN 1028-6020. [CrossRef] [PubMed].
27. Arif T, Mandal TK, Dabur R. "Natural products: Anti-fungal agents from plants". In: Opportunity, Challenge and Scope of Natural Products in Medicinal Chemistry; V.K. Tiwari, B.B. Mishra (editors), Research Signpost: Kerala, India, 2011; pp. 283-311. ISBN 9788130804484.
28. Mansfield BE, Oltean HN, Oliver BG, Hoot SJ, Leyde SE, Hedstrom L et al. Azole drugs are imported by facilitated diffusion in Candida albicans and other pathogenic fungi. PLoS Pathog. 2010; 6(9). [CrossRef].
29. Bard M, Lees ND, Turi T, Craft D, Cofrin L, Barbuch R et al. Sterol synthesis and viability of erg11 (cytochrome P450 lanosterol demethylase) mutations in Saccharomyces cerevisiae and Candida albicans. Lipids. 1993; 28(11): 963-967. ISSN 1558-9307. [CrossRef] [PubMed].
30. Lamb DC, Kelly DE, Schunk WH, Shyadehi AZ, Akhtar M, Lowe DJ et al. The mutation T315A in Candida albicans sterol 14alpha-demethylase causes reduced enzyme activity and fluconazole resistance through reduced affinity. J Biol Chem. 1997; 272(9): 5682-5688. ISSN 1083-351X. [CrossRef] [PubMed].
31. Irfan M, Abid M. Three dimensional structure modeling of lanosterol 14-α demethylase of Candida albicans and docking studies with new triazole derivatives. Chem inform. 2015; 1(1,4):1-8. ISSN 2470-6973. [Link].
32. Bhatnagar RS, Fütterer K, Farazi TA, Korolev S, Murray CL, Jackson-Machelski E et al. Structure of N-myristoyl-transferase with bound myristoylCoA and peptide substrate analogs. Nat Struct Biol. 1998; 5(12): 1091-1097. ISSN 1072-8368. [CrossRef] [PubMed].
33. Gelb MH, Van Voorhis WC, Buckner FS, Yokoyama K, Eastman R, Carpenter EP etal. Protein farnesyl and N-myristoyl transferases: Piggy-back medicinal chemistry targets for the development of antitrypanosomatid and antimalarial therapeutics. Mol Biochem Parasitol. 2003; 126(2):155-63. ISSN 0166-6851. [CrossRef] [PubMed].
34. Wright MH, Heal WP, Mann DJ. Tate EW. Protein myristoylation in health and disease. J Chem Biol. 2010; 3(1): 19-35. ISSN 1864-6158. [CrossRef] [PubMed].
35. Guerrero-Perilla C, Bernal FA, Coy-Barrera ED. Molecular docking study of naturally-occurring compounds as inhibitors of N-myristoyl transferase towards antifungal agents discovery. Rev Colomb Cien Quím Farm. 2015; 44(2): 162-178. ISSN 0034-7418. [CrossRef].
36. Wakelin SA, Ryder MH, Warren RA. Effect of soil properties on growth promotion of wheat by Penicillium radicum. Aust J Soil Res. 2004; 42(8): 897–904. ISSN 0004-9573. [CrossRef].
37. Vining LC. Functions of secondary metbolites. Annu Rev Microbiol. 1990; 44: 395-427. ISSN 0066-4227. [CrossRef].
38. Singh V, Tripathi CKM, Katti SB, Praveen V, Tripathi D, Haque S. Isolation, characterization and antifungal docking studies of wortmannin isolated. Sci Rep. 2015; 5: 11948. [CrossRef].
39. Arendrup MC, Fuursted K, Gahrn-Hansen B, Jensen IM, Knudsen JD, Lundgren B et al. Seminational surveillance of fungemia in Denmark: notably high rates of fungemia and numbers of isolates with reduced azole susceptibility. J Clin Microbiol. 2005; 43(9): 4434-4440. ISSN 0095-1137. [CrossRef] [PubMed].
40. Teixeira MM, Theodoro RC, de Carvalho MJ, Fernandes L, Paes HC, Hahn RC et al. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol Phylogenet Evol. 2009; 52(2): 273-283. ISSN 1095-9513. [CrossRef] [PubMed].
41. Restrepo A, Benard G, Castro CC, Agudelo CA, Tobón AM. Pulmonary paracoccidioidomycosis. Semin Respir Crit Care Med. 2008; 29(2): 182-197. ISSN 1069-3424. [CrossRef] [PubMed].
42. Abadio AKR, Kioshima ES, Leroux V, Martins NF, Maigret B, Felipe MSS. Identification of new antifungal compounds targeting thioredoxin reductase of Paracoccidioides Genus. PLoS ONE. 2015; 10(11): e0142926. [CrossRef].
43. Ghannoumand MA, Rice LB. Antifungal agents: mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin Microbiol Rev. 1999; 12(4): 501-517. ISSN 1098-6618. [PubMed].
44. Arendrup MC, Patterson TF. Multidrug-resistant Candida: epidemiology, molecular mechanisms, and treatment. J Infect Dis. 2017; 216(3): S445-S451. ISSN 1537-6613. [CrossRef].
45. Edlind TD, Henry KW, Metera KA, Katiyar SK. Aspergillus fumigatus CYP51 sequence: potential basis for fluconazole resistance. Med Mycol. 2001; 39(3): 299-302. ISSN 1369-3786. [CrossRef] [PubMed].
46. Chowdhary A, Prakash A, Sharma C, Kordalewska M, Kumar A, Sarma S et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009–17) in India: role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J Antimicrob Chemother. 2018; 73(4): 891-899. ISSN 1460-2091. [CrossRef] [PubMed].
47. Chowdhary A, Sharma C, Meis JF. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017; 13(5): 1-10, e1006290. [CrossRef].
48. Lockhart SR, Etienne KA, Vallabhaneni S, Farooqi J, Chowdhary A, Govender NP et al. Simultaneous emergence of multidrug-resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin Infect Dis. 2017; 64(2):134-140. ISSN 1537-6591. [CrossRef] [PubMed].
49. Clinical Alert to U.S. Healthcare Facilities—June 2016. Global Emergence of Invasive Infections Caused by the Multidrug-Resistant Yeast Candida auris. [Link].
50. Candida auris in Healthcare Settings-Europe. [Link].
51. Pan American Health Organization/ World Health Organization (PAHO/ WHO). Candida auris' Outbreaks in Health Care Services — Epidemiological Alert. [Link].
52. Leonardelli F, Macedo D, Dudiuk C, Cabeza MS, Gamarra S, Garcia-Effron G. Aspergillus fumigatus intrinsic fluconazole resistance is due to the naturally occurring T301I substitution in Cyp51Ap. Antimicrob Agents Chemother. 2016; 60(9): 5420-5426. ISSN 1098-6596. [CrossRef] [PubMed].
53. Doligalski CT, Benedict K, Cleveland AA, Park B, Derado G, Pappas PG et al.Epidemiology of invasive mold infections in lung transplant recipients. Am J Transplant. 2014; 14(6): 1328-1333. ISSN 1600-6143. [CrossRef] [PubMed].
54. Steinbach WJ, Marr KA, Anaissie EJ, Azie N, Quan SP, Meier-Kriesche HU et al. Clinical epidemiology of 960 patients with invasive aspergillosis from the PATH Alliance registry. J Infect. 2012, 65(5): 453-464. ISSN 1532-2742. [CrossRef] [PubMed].
55. Garcia-Effron G, Dilger A, Alcazar-Fuoli L, Park S, Mellado E, Perlin DS. Rapid detection of triazole antifungal resistance in Aspergillus fumigates. J Clin Microbiol. 2008; 46(4): 1200-1206. ISSN 1098-660X. [CrossRef] [PubMed].
56. Walsh TJ, Anaissie EJ, Denning DW, Herbrecht R, Kontoyiannis DP, Marr KA et al. Treatment of aspergillosis: clinical practice guidelines of the Infectious Diseases Society of America. Clin Infect Dis. 2008; 46(3): 327-360. ISSN 1537-6591. [CrossRef] [PubMed].
57. Prasad R, Sharma M, Kaur Rawal M. Functionally Relevant Residues of Cdr1p: A Multidrug ABC Transporter of Human Pathogenic Candida albicans. J Amino Acids, 2011, Article ID 531412. ISSN 2090-0112. [CrossRef].
58. Sanglard D, Ischer F, Monod M, Bille J. Cloning of Candida albicans genes conferring resistance to azole antifungal agents: characterization of CDR2, a new multidrug ABC transporter gene. Microbiology. 1997; 143(Pt2): 405-416. ISSN 1465-2080. [CrossRef] [PubMed].
59. Prasad R, Balzi E, Banerjee A, Khandelwal NK. All about CDR transporters: Past, present, and future. Yeast. 2019; 36(4): 223-233. [CrossRef].
60. Chen Z, Li J, Wang W, Guo X, Li Y, Mao X et al. Mutations adjacent to the end of transmembrane helices 6 and 7 independently affect drug efflux capacity of yeast ABC transporter Pdr5p. Biochim Biophys Acta. 2014; 1838(3): 932-939. ISSN 0005-2736. [CrossRef].
61. Nim S, Monico A, Rawal MK, Duarte N, Prasad R, Di Pietro A et al. Overcoming multidrug resistance in Candida albicans: Macrocyclic diterpenes from Euphorbia species as potent inhibitors of drug efflux pumps. Pl Med. 2016; 82(13): 1180-1185. ISSN 0032-0943. [CrossRef] [PubMed].
62. Puri N, Prakash O, Manoharlal R, Sharma M, Ghosh I, Prasad R. Analysis of physico-chemical properties of substrates of ABC and MFS multidrug transporters of pathogenic Candida albicans. Eur J Med Chem. 2010; 45(11): 4813-4826. ISSN 0223-5234. [CrossRef].
63. Rutledge RM, Esser L, Ma J, Xia D. Toward understanding the mechanism of action of the yeast multidrug resistance transporter Pdr5p: A molecular modeling study. J Struct Biol. 2011: 173(2): 333-344. ISSN 1047-8477. [CrossRef] [PubMed].
64. Diniz WJS, Canduri F. Review article. Bioinformatics: an overview and its applications. Genet Mol Res. 2017; 16(1): 1-21. ISSN 1676-5680. [Link]. [CrossRef].
65. Bencurova E, Gupta SK, Sarukhanyan E, Dandekar T. Review. Identification of antifungal targets based on computer modeling J Fungi (Basel). 2018; 4(3): 81. ISSN 2309-608X. [CrossRef].
66. Chaskar P, Zoete V, Röhrig UF. Toward On-The-Fly Quantum Mechanical/Molecular Mechanical (QM/MM) Docking: Development and Benchmark of a Scoring Function. J Chem Inf Model. 2014; 54(11): 3137−3152. ISSN 1520-5142.
67. Chen SJ. A potential target of Tanshinone IIA for acute promyelocytic leukemia revealed by inverse docking and drug repurposing. Asian Pac J Cancer Prev. 2014; 15(10): 4301-4305. ISSN 2476-762X. [CrossRef] [PubMed].
68. Singh VK, Singh AK, Chand R, Kushwaha C. Role of Bioinformatics in agriculture and sustainable development. Int J Bioinformatics Res. 2011; 3(2): 221-226. ISSN 0975-9115. [Link].
69. Taylor WR, Orengo CA. Protein structure alignment. J Mol Biol. 1989; 208(1): 1-22. ISSN 0022-2836.
70. Lipman DJ, Altschul SF, Kececioglu JD. A tool for multiple sequence alignment. Proc Natl Acad Sci USA. 1989; 86(12): 4412-4415. ISSN 1091-6490 .